Chapter 3 Morphology of the outer ear
INTRODUCTION
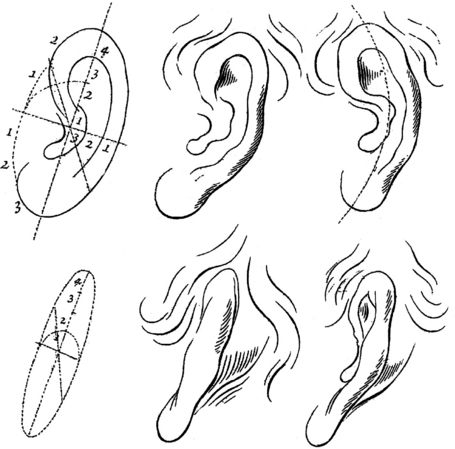
The shape of the outer ear has long been a subject of interest to both artists and anthropologists. Artists have been impressed by the harmony of its lines and curves and much study was needed before they were able to reproduce it in paintings and drawings. For learners, a geometric scheme of the ear imagined to be inscribed in an ellipse was proposed in 1768.1 The upper drawing in Figure 3.1 represents a major semi-axis divided into four parts and a minor semi-axis (half of the former) divided into three parts. The diagonal axis subdivides the upper left quarter into two approximately equal parts and this line is intended to indicate the prominent point of the helix at the junction between the first and second part. The lower left quarter is subdivided into three parts to localize the prominent point of the ear lobe at the junction between the second and third part (Fig. 3.1).
THE ORGANIC EVOLUTION OF THE PINNA IN PRIMATES AND HUMANS
Charles Darwin (1809–1882), in his masterpiece The Descent of Man, and Selection in Relation to Sex,2 wrote:
The whole external shell of the ear may be considered a rudiment, together with the various folds and prominences (helix and anti-helix, tragus and anti-tragus &c.) which in the lower animals strengthen and support the ear when erect, without adding much to its weight …
The celebrated sculptor, Mr. Woolner, informs me of one little peculiarity in the external ear, which he has often observed both in men and women, and of which he perceived the full signification. The peculiarity consists in a little blunt point, projecting from the inwardly folded margin or helix. These points not only project inwards, but often a little outwards, so that they are visible when the head is viewed from directly in front or behind. They are variable in size and somewhat in position, standing either a little higher or lower; and they sometimes occur on one ear and not on the other … we may safely conclude that it is a similar structure – a vestige of formerly pointed ears – which occasionally reappears in man. (Fig. 3.2)
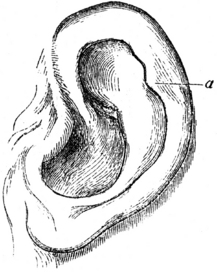
Fig. 3.2 Human ear, modeled and drawn by the sculptor Mr Woolner.2 Point marked a = the ‘projecting point’ mentioned by Darwin.
Darwin2 made several comparative observations in primates and could not at his time understand the reason of the loss of mobility of the outer ear. He wrote:
Darwin’s observations influenced all following anthropologists who tried to find an answer to this query. Two famous researchers, the German Gustav Schwalbe (1844–1916) and Rudolf Martin (1864–1925) from Switzerland, demonstrated that the process of regression in primates and in man consisted essentially in a shortening of the tip of the ear, causing an inward curl of the helix and a rise of the upper branch of the anthelix. Schwalbe3 measured the base of the ear (a–b in his study) and the so-called true length (d–c), from the tragus to Darwin’s tubercle, and compared the morphological ear index (base × 100/true length) in various mammals and primates (Fig. 3.3; Box 3.1). Humans had the highest score not only because length was reduced but also because the base of the ear was significantly larger. This increase was actually responsible for the progressive loss of mobility observed by Darwin. Nevertheless, the significance of the raised anthelix and the curled helix remained obscure and it was only several decades later that an explanation was found, thanks to modern technology.
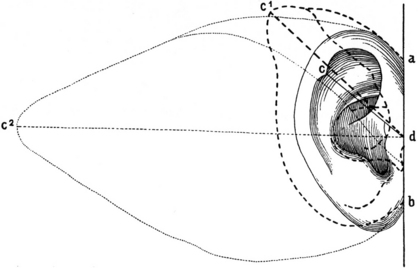
Fig. 3.3 The human outer ear compared with that of a baboon and that of a calf according to Schwalbe.3 The human outer ear (represented by an unbroken line; a c b); the outer ear of a baboon (dashed line; a c1 b) and the outer ear of a calf (dotted line; a c2 b) were overlapped maintaining the same base length (a d b).
Box 3.1 The Morphological Ear Index measured in different mammals: the higher scores belong to primates and humans, according to Schwalbe3

The unanswered questions in recent times were fundamentally two:
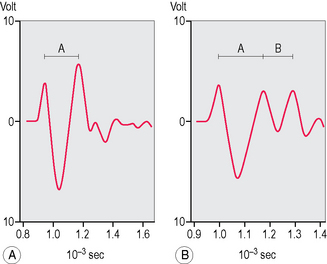
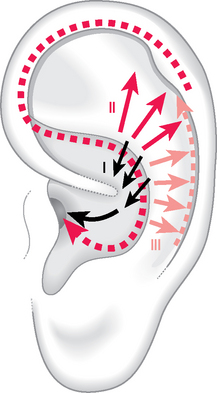
Several hypotheses were proposed on both subjects but the most interesting conclusions were probably those reached by the psychiatrist Johann Burchard4–6 of Hamburg, who dedicated 30 years of his life to these issues. In a series of experiments he noticed that a square-shaped click of 0.14 ms produced by the computer could be recorded with two or three peaks by a miniaturized microphone in the external acoustic meatus (Fig. 3.4A, B). Of 53 people (106 ears) placed in a noiseless room, 14% showed two peaks and 86% three peaks. In both groups interval A (between the first and second peak) and interval B (between the second and third peak) were significantly different when the direction of the sound was shifted from a perpendicular axis to the ear surface to points placed at 45° respectively in front, at the back and below (P <0.01) or up (P <0.05). These data confirmed the evidence that the human ear can actually localize sounds in space without moving the pinna.
Burchard’s second hypothesis was that the three peaks could correspond to three different anatomical paths covered by the sound wave transmitted to the external meatus (Fig. 3.5). Calculating the average delays from one peak to the next, he obtained distances in cm which were significantly correlated with the length of the respective anatomical path measured with soft threads (P <0.01). He identified three possible paths: the first (I in Fig. 3.5) was the shortest way to reach the external meatus (about 0.1 ms corresponding to 3 cm) and could seemingly start from the lower concha and the wall of the anthelix; the second (II in Fig. 3.5) had a further average delay of 6.6 cm and could possibly start from the upper-middle portion of the scapha and helix tunnel; the third (III in Fig. 3.5) had a further average delay of 3 cm and could possibly originate from the lower portion of the same anatomical parts. The incomplete consistency of peak 3 was explained by Burchard by the lack in these subjects of a sufficiently pronounced depression of this anatomical portion (as for example in Fig. 3.6 or in types 3 and 4 in Fig. 3.9A). Repeating the same experiment on chimpanzees and gorillas he found that these primates had only two peaks which were similar to peaks 1 and 2 in humans. The less curled helix and the distance between anthelix and helix make the third peak, typical in humans, impossible for primates. The author concluded that the triple signal and its variations of amplitude and interval were essential for the cortical recognition of sound which is fundamental in the analysis of speech.
ANTHROPOMETRY OF THE OUTER EAR
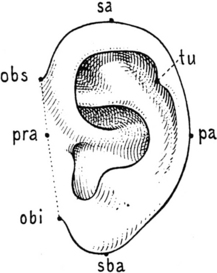
Anthropometrists, dealing with the measurement of size, weight and proportions of the human body, also introduced some anatomical landmarks for the outer ear to be used for calculating its dimension. These are otobasion superius (obs), otobasion inferius (obi), which are the upper and lower points at which the pinna is attached to the scalp (Fig. 3.7). The line joining obs and obi is called the base of the ear. The other points are superaurale (sa) and subaurale (sba), which allow the measurement of the so-called ‘physiognomic length’, and preaurale (pra) and postaurale (pa), which are used to measure the ‘physiognomic breadth’ of the pinna. Both measurements are usually taken by the anthropologist using a special calliper.3 Among the data gathered by different authors on large samples of the population are the average larger dimensions of the pinna in males compared to females. Another interesting phenomenon observed by various researchers is the constant increase of both measurements, particularly of length, during a subject’s life.3,7 Especially after the age of 50 there is a further increase which has to be correlated with various factors such as the flattening of the anthelix and the reduction of curling of the helix, but also the decrease of skin elasticity and the particular growth of the ear lobe. In addition the appearance of creases in the front of the tragus and the growth of hair on the external meatus are considered good parameters for judging a subject’s age.
MORPHOLOGICAL VARIABLES
In the past anthropologists have studied the outer ear to make a reliable classification of the dominant and recessive trait of some recurrent morphological characteristics. Studies have been carried out on large groups of individuals of different racial and geographical origin. Another application of the inspection of the outer ear, nowadays superseded by genomic analysis, was sometimes proposed by forensic medicine as a control in the scientific clarification of paternity.8,9
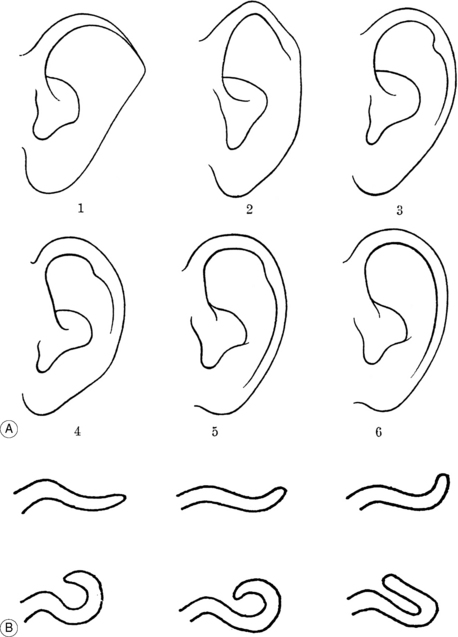
The effort made by anthropologists to describe the morphological variables of the ear as exhaustively as possible is of great value and interest to us. The reader could actually be helped to differentiate some morphological characteristics with a more pronounced hereditary tendency than others acquired later in the patient’s life (see the next section). Figure 3.8 reports the first which received particular consideration in the literature as a possible dominant or recessive trait.7 In this description I have excluded any malformations associated with hereditary deafness, renal and vertebral anomalies and chromosome abnormalities such as trisomy 18 syndrome. The frequency of the illustrated characteristics is very variable: for example, according to Lange, in 908 subjects: the curled helix was visible in 60.1%; Darwin’s tubercle in 59.8%; the tuberculum supratragicum in 38%; the helix taeniata in 9%; the crus cymbae in 6.8%; the incisura prelobularis in 4.7%; the crus anthelicis tertium in 3.4%. Some significant sexual differences were observed, among them the higher frequency of Darwin’s tubercle in males and the strikingly higher proportion of curled helix in females (76.8% in females compared with 44.2% in males)3 (Fig. 3.9). These characteristics are seemingly correlated and Martin3 gives us the following explanation to think about, and perhaps be amused by:
< div class='tao-gold-member'>
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


