CHAPTER 4 Transduction, Transmission and Perception of Pain
Painful spinal disorders are common problems in society, affecting an estimated 50 million Americans. The societal costs (including litigation, work lost, treatment, and disability) for such disorders of the spine are staggering. For example, the cost of low back pain alone has been estimated at US$40–50 billion annually.1,2 Chronic neck pain has a similarly high cost of nearly US$30 billion in health-related expenses.3 Until a better understanding of the pathomechanisms of pain and the injuries which produce them are defined, the effective prevention and treatment of these disorders and their symptoms will remain elusive. Further, distinguishing those physiologic mechanisms which lead to persistent pain from those which differentially produce only transient symptoms is also important in understanding and managing these syndromes. It is the intent of this chapter to highlight traditional and emerging theories of pain detection and transmission in the context of spine-related syndromes. A brief discussion of the neurophysiology of pain highlights concepts of local responses, pain transduction, signal transmission, and processing and is integrated with more recent hypotheses of the central nervous system’s (CNS) neuroimmunologic involvement in persistent pain.
It is important to define, at the outset, relevant distinctions in terminology. ‘Pain’ is a complex perception that is influenced by prior experience and by the context within which the noxious stimulus occurs. Likewise, ‘nociception’ is the physiologic response to tissue damage or prior tissue damage. Similarly, for discussion in this chapter, ‘hyperalgesia’ is defined as enhanced pain to a noxious stimulus.4 Strictly speaking, this is a leftward shift of the stimulus–response function relating pain to intensity. The corresponding pain threshold is lowered and there is enhanced response to a given stimulus. Hyperalgesia is mediated by nociceptor sensitization, where ‘sensitization’ describes a corresponding shift in the neural response curve for stimulation. Sensitization is characterized by a decrease in threshold, an increased response to suprathreshold stimulus, and spontaneous neural activity.
RELEVANT NEURAL ANATOMY
Before presenting and discussing pain mechanisms, it is first necessary to describe the relevant anatomical structures, biological connections, and relationships of neural sensory and processing components. These are reviewed only briefly here to provide appropriate context; a more detailed presentation can be found in texts specifically focused in neural science and pain.4,5
The primary afferents, which relay pain signals from injured or stimulated tissues, terminate in the dorsal horn of the spinal cord. At each level in the spinal cord, the dorsal nerve roots carry sensory information from the periphery into the spinal cord. Dorsal roots contain sensory neurons, whose cell bodies make up the enlarged dorsal root ganglion (DRG) (Fig. 4.1). In contrast, the ventral root contains the axons of neurons whose cell bodies are within the ventral horn of the spinal cord and transmits efferent signals. At each spinal level, the dorsal and ventral nerve roots come together, outside of the spinal column and distal to the DRG, and combine to form the nerve which communicates with the peripheral nervous system. The spinal nerves further branch into smaller nerves in the periphery and innervate bones, ligaments, joints, discs, muscles, organs, and many other tissue types.
Structurally, three protective layers surround the spinal cord, which are themselves extensions of the cranial meninges: the dura mater (outermost), the arachnoid mater, and the pia mater (innermost layer closest to the spinal cord). Within the spinal column, the lumbar dorsal and ventral nerve roots extend below the spinal cord and this neural tissue, collectively called cauda equina, fills the sacral spinal column. The spinal cord is anatomically composed of two regions (Fig. 4.2). These are distinguished by their appearance, function, and cell populations. The gray matter, which has a darker appearance, contains the cell bodies of spinal neurons and makes up the central region of the spinal cord. It is surrounded by the white matter which contains the axons of the spinal neurons. The columnar tracts of the spinal cord are regionally specialized according to information they carry (see Fig. 4.2). The lateral column contains motor neurons; the dorsal column carries information related to mechanoreception; and the ventrolateral column houses neurons which communicate information regarding pain, temperature, and motor signals. In general, the sensory system ascending pathway comprises the dorsal portion of the spinal cord, while the descending pathway of the motor control system comprises the ventral aspect of the cord.
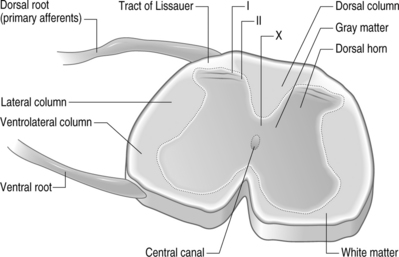
Afferents of the dorsal nerve root enter the spinal cord dorsolaterally and branch in the white matter, with collaterals which terminate in the gray matter. Nerve fibers mediating pain pass through the tract of Lissauer and have branches which terminate in the most superficial regions of the dorsal horn, laminae I and II. Neurons in these laminae synapse on secondary neurons in laminae IV–VI of the dorsal horn and these secondary neurons cross the midline before ascending to the brain contralaterally in the anterolateral region of the cord. Lamina X, which is located in the gray matter region closest to the central canal, also receives sensory inputs related to pain. The neurons of the substantia gelatinosa receive information from Aδ and C fibers; Aβ afferents terminate in the deeper laminae. After injury, it is believed that Aβ afferents sprout from the deeper lamina into the dorsal horn where they make synaptic contacts with neurons.6,7
Each regional level of the spinal cord receives sensory information from specific regions of the body, known as dermatomes. Typically, nerves from approximately two spinal levels innervate any given region of the skin’s surface. These surfaces have been divided into discrete regions, providing a dermatomal map relating each region of the skin to a corresponding spinal level.8 Clinically, dermatomal maps are used to identify the origin of painful symptoms. However, nerve endings which innervate internal organs can also produce cutaneous sensation. This ‘referred pain’ sensation is experienced at sites other than its source and is due to the fact that nearly all spinal neurons that innervate internal organs also are associated with cutaneous sensation.
TRANSDUCTION
Nociceptive afferents are specific for sensing different noxious stimuli: thermal, mechanical, and chemical stimuli. Some nociceptors are polymodal and sense all types of stimuli. Broadly, sensory nerve fibers range in diameter from <0.05 μm to 20 μm. These fibers are either unmyelinated or myelated with thick or thin myelin sheathes enclosing them. Their conduction velocities can range from 0.5 m/s to 120 m/s, depending on their axon diameter and/or the presence of myelination. A fibers are myelinated and can evoke sharp and pricking pain sensation; they can also convey sensations of aching pain. The largest, myelinated sensory axons, Aβ, are generally classified as mechanoreceptive. Aβ fibers are primarily proprioceptive (sensing mechanical movement in joints and muscles). Their diameters are approximately 10 μm; these fibers have slower conduction velocities than Aδ fibers. The smallest myelinated fibers are Aγ fibers, which primarily mediate pin-prick, itching, and other mechanical sensations of pain. Unmyelinated C fibers mediate thermal sensation, in addition to mechanical pain. Stimulation of C fibers can also induce a burning sensation. C and some A fibers are the primary high-threshold noceiceptors. The A fibers exhibit greater response frequencies than C fibers and have more communications to the spinal cord.
Pain detection and signaling begin at the injury site where Aδ and C fibers are activated by thermal, chemical, mechanical, and/or electrical stimuli. These nociceptors can become sensitized, lowering their thresholds for firing and increasing their firing rates when stimulated.9 Further, damaged cells at the site of injury release ATP and free protons. Briefly, a complex series of reactions also occur, including altered blood flow, release of neuropeptides from the afferent fiber itself, and a generalized host of biochemical and cellular responses typical of an immune response. For painful tissue injuries, inflammation is induced in an effort to promote healing and recovery. In this process, inflammatory mediators such as prostaglandin E, serotonin, bradykinin, and histamine, among others, can alter fiber responses. These and other mediators directly activate nociceptors or can sensitize those nociceptors which are already responding to stimuli. This further induces increased neuronal activity. Specifically, bradykinin, serotonin, excitatory amino acids, and hydrogen ions are all responsible for directly activating afferents.9,10 Similarly, prostaglandins, serotonin, noradrenaline, adenosine, nitric oxide, and nerve growth factors can sensitize nociceptors.
In addition to altered electrical activity, and the local synthesis and release of inflammatory mediators that induce inflammation and edema as part of the healing process, these same immune processes that provide healing also sensitize nociceptors and recruit new nociceptors that enhance pain.11,12 In particular, cytokines are released in the periphery in association with tissue injury and inflammation. These small proteins, in turn, contribute to the local inflammatory response, while further modulating electrophysiologic responses of nerve fibers and altering nociception. Additional details regarding specific biochemical mediators of pain and their mechanisms of action throughout the pain signaling pathway (i.e. injury site and spinal cord) are provided in a later section specifically addressing biochemical mediators of pain.
TRANSMISSION
In general, injury to a variety of tissues, including muscle, disc, ligament, and neural tissue, can induce many cascades and physiological signals which lead to pain transmission (Fig. 4.3). During this process, neuroplasticity and subsequent CNS sensitization responses produce altered functions of chemical, electrophysiological, and pharmacological systems throughout the periphery and central systems.4,13–17 This cascade of biochemical and molecular reactions causes changes in gene transcription, post-translational modifications, and culminates with the transmission of pain.18 The interplay of these system responses involves complicated cross-system effects between injury and changes in both the peripheral and central nervous systems. The integration of multiple physiologic systems, as occurs with pain transmission (see Fig. 4.3), contributes to the overall challenges in preventing such syndromes since a given insult may initiate a host of different responses which can be established and maintained remote from the actual site of injury. Regardless, there is a generalized series of responses which occurs following a painful injury in the periphery.
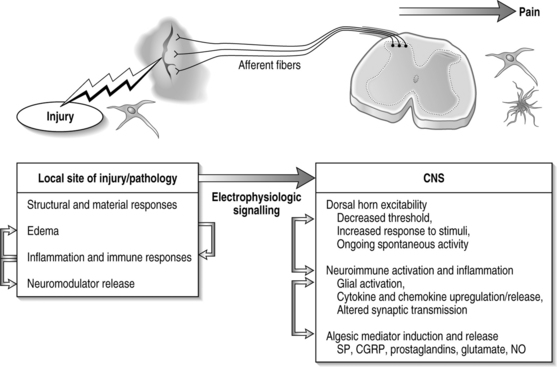
A chemical insult in the periphery causes direct activation of sensory neurons; voltage-gated sodium channels are integral for initiating and propagating pain signals in these sensory neurons. Two separate types of such channels have been designated as SNS and SNS-2, respectively. The SNS subtype is present on both C and some A fibers; SNS-2 channels are exclusively present on unmyelinated neurons. The vanilloid receptor (VR1) is spatially co-localized with these channels.
Calcium concentrations play a critical role in mediating pain in the central nervous system by controlling and regulating levels of nitric oxide synthase (NOS), prostaglandins, and changes in their gene transcription.19 An increase in calcium causes enhanced release of neurotransmitters, as well as an increase in activity of neuronal nitric oxide synthase. Because changes in intracellular calcium occur through the actions of several different channel types, it is possible to isolate the effects of calcium based on which channel opened. For example, in striatal neurons, the population of calcium that activates nNOS is derived from opening of a voltage-gated calcium channel. In addition, increases in intracellular calcium via NMDA receptor influx causes increases in arachidonic acid which can enable COX and prostaglandin production. This process sensitizes neurons of the dorsal horn by a protein kinase C pathway; this second messenger can induce neurons to release substance P and glutamate and provide nociceptive transmission.
Primary afferents communicate with spinal neurons via synaptic transmission. A variety of neurotransmitters (i.e. glutamate, NMDA, substance P) modulate postsynaptic responses, with further transmission to postsynaptic spinal neurons and supraspinal sites via the ascending pathways.9 Peripheral signals of injury pain generate increased neuronal excitability in the spinal cord.17 Associated with this sensitization are a decreased activation threshold, increased response magnitude, and increased recruitment of receptive fields.4 The continuous input from nociceptive afferents can drive spinal circuits and lead to central sensitization, maintaining a chronic pain state.15 These neuroplastic changes are accompanied by other electrophysiological manifestations that cause neurons to fire with increased frequency and even to fire spontaneously.16 Spinal processing is further directly altered by descending inhibitory and facilitory pathways that can provide additional modulation of spinal interneurons.20
Ultimately, persistent pain results from the sensitization of the central nervous system. While the exact mechanism by which the spinal cord reaches a ‘hyperexcitable’ state of sensitization is not fully known, many hypotheses have emerged. Here, only highlights of these theories are provided as an overview. More extensive and detailed discussions can be found elsewhere in the literature.17,21–23 Central hyperexcitability is characterized by a ‘wind-up’ response of repetitive C fiber stimulation, expanding receptive field areas, and spinal neurons taking on properties of wide dynamic-range neurons.24 Low threshold Aβ afferents, which normally do not transmit pain, begin to signal spontaneous and movement-induced pain.22 Aβ fibers stimulate postsynaptic neurons to transmit pain, where these Aβ fibers previously had no effect. This plasticity of neuronal function in the spinal cord contributes to central sensitization.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


