TABLE 61.1 Characteristics of the Three Metabolic Systems | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||
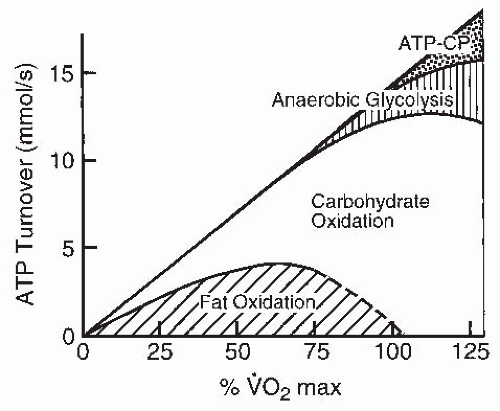 FIGURE 61-1. Relative importance of the different metabolic systems as a function of exercise intensity. (Adapted from Sahlin K. Metabolic changes limiting muscle performance. In: Saltin B, ed. Biochemistry of Exercise VI. Champaign, IL: Human Kinetics; 1986;323-343.) |
diet, mode of exercise, and level of fitness. As the intensity of the exercise is increased, the predominant fuel source shifts toward carbohydrate. This is partly because ATP production shifts toward anaerobic metabolism during high-intensity exercise and carbohydrate is the only fuel available for anaerobic glycolysis. Carbohydrates are made available to the contracting muscle through mobilization of muscle and liver glycogen stores as well as through ingested carbohydrates that are circulating in the bloodstream.
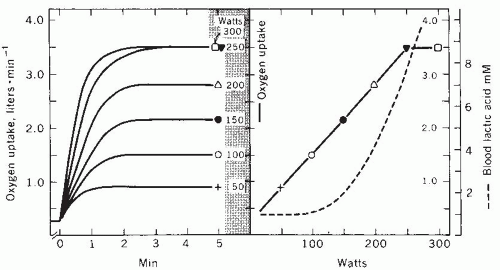 FIGURE 61-2. Schematic demonstration of the determination of maximal oxygen uptake. Left: Oxygen uptake increases during 5-minute exercise stages on a bicycle ergometer at different workloads (noted within the shadowed area). Right: Oxygen uptake at each workload measured after 5 minutes, plotted in relation to workload. Note that there was no additional increase in oxygen uptake between the 250- and the 300-W workloads. Maximal oxygen uptake is 3.5 L/min. Blood lactic acid concentrations across each workload are also demonstrated. (With permission from Astrand PO, Rodahl K. Textbook of Work Physiology: Physiological Basis of Exercise. 3rd ed. New York: McGraw-Hill; 1986.) |
TABLE 61.2 Characteristics of the Different Metabolic Substrates | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
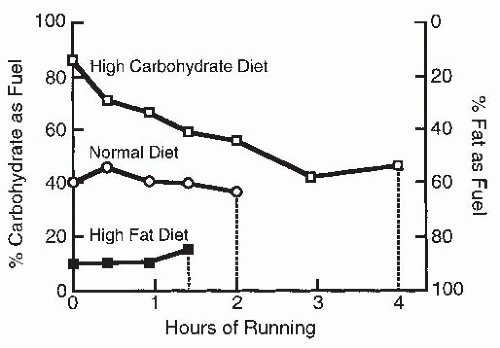 FIGURE 61-3. Effect of three different diets on fuel source used during running. Vertical bars represent the points at which exhaustion occurred. (Based on data from Christensen EH, Hansen O. Arbeitsfahigkeit und Ermahrung. Skand Arch Physiol. 1939;8:160-175.) |
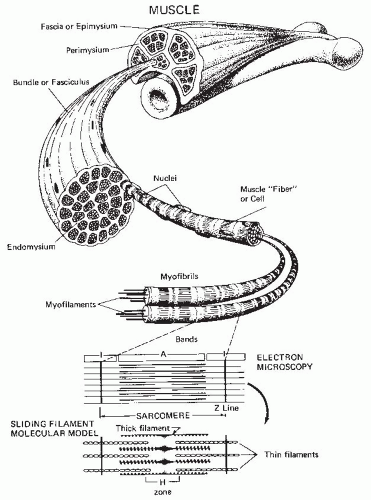 FIGURE 61-4. Structural and functional subunits of skeletal muscle. (With permission from Lamb DR. Physiology of Exercise: Responses and Adaptations. 2nd ed. New York: Macmillan; 1984.) |
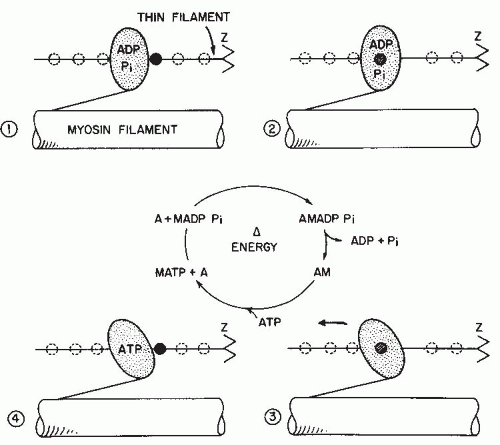 FIGURE 61-5. Postulated mechanism of crossbridge formation and cycling by actin and myosin filaments. The head of the myosin filament is stippled and forms the crossbridge between the two filaments. Active sites on the actin (thin) filament are outlined by broken circles. 1: No bonds between filaments. 2: Initial attachment of myosin head to one of the active sites on the actin filament. This attachment takes place only in the presence of Ca2+ ions. 3: Formation of strong actin-myosin bond. This process causes a conformational change in the angle of the myosin head, which produces a relative movement of actin and myosin filaments across one another. ADP and phosphate ions are lost from the myosin. 4: If ATP is available, the actin-myosin bond can be broken. Subsequent hydrolysis of the ATP by the myosin ATPase returns the cycle to stage 1. If no ATP is available, the actin-myosin bond remains intact, as in muscular rigor. (From Gordon AM. Muscle. In: Ruch T, Patton H, eds. Physiology and Biophysics. Vol. IV. Philadelphia, PA: WB Saunder; 1982:170-260. Reprinted by permission of WB Saunders Co.) |
contractile element with both series and parallel elastic elements. The contractile element actively generates force and represents the interaction between actin and myosin filaments. The elastic elements are purely passive components acting as mechanical springs. The series elastic element represents the tendinous insertions of muscle, and the parallel elastic element represents the connective tissue surrounding the various subunits of the muscle.
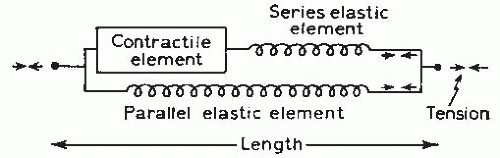 FIGURE 61-6. Mechanical model of muscle consisting of a contractile element and two elastic elements. (From Roberts TDM. Neurophysiology of Postural Mechanics. London: Butterworth; 1978.) |
TABLE 61.3 Muscle Fiber Type Continuum Using the Myosin ATPase Classification System | |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
phosphorylase. Phosphorylase is an enzyme that is involved in the breakdown of glycogen. As a result, these fibers are suited for performance of high-intensity, short-duration work. Hybrid subtypes exist linking the major fibers of I and IA and IX on a continuum from type I to IC to type IIC, IIAC to IIA to IIAX to IIX. With activation of the motor units, a transition of the ATPase isoforms will move from the type IIX to the type IIA subtype (17). The continuum of hybrids fibers goes from the most oxidative isoform type I to the least oxidative isoform type IIX. However, again, when a motor unit is activated, oxidative processes increase and exercise training will even leave type IIX fibers that remain with higher concentrations of oxidative enzymes. Thus, the transition to the primary fiber type as visualized by the histochemical staining is related to the oxidative status of the fiber while it keeps its characteristic size and function. Capillary development also follows this transition to greater density as oxidative needs increase. However, due to the fact that type I motor units are orderly recruited first in almost all activities and are involved for the repetitive endurance activities, their oxidative, mitochondrial, and capillary profiles are always greater than type II or fast motor units.
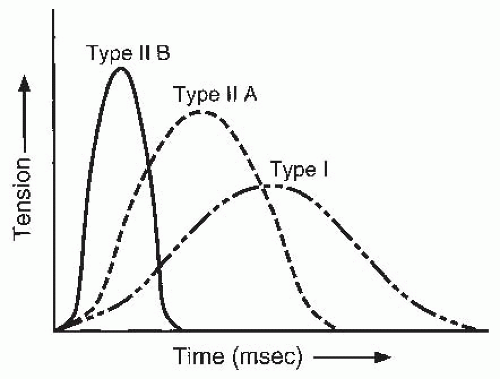 FIGURE 61-7. Schematic representation of rate of tension generation and force production with different fiber types. The speed of contraction and force are greatest for type II fibers. |
inhibition of the protective reflex mechanisms. Furthermore, the bilateral deficit has been shown to be reduced through training with bilateral contractions (33).
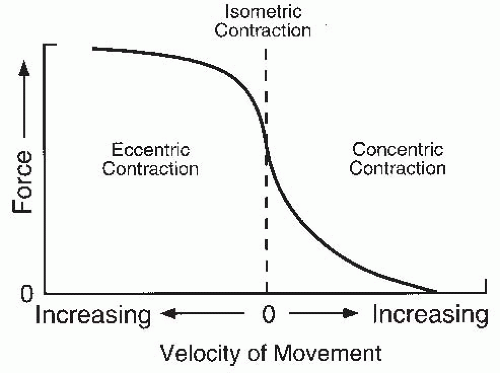 FIGURE 61-8. Schematic relationship between maximal muscular force and velocity of movement. |
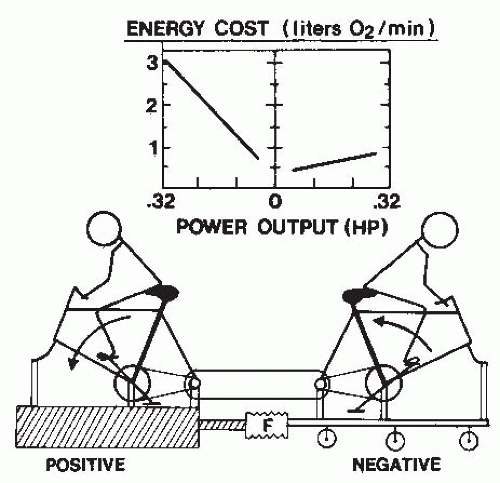 FIGURE 61-9. Demonstration of the difference in energy cost between positive and negative work. The two subjects pedaled coupled bicycles with one pedaling forward (performing positive work) and the other providing the resistance as the cranks rotated backward (performing an equal amount of negative work). Graph inset shows the differences in energy costs across the examined power outputs. (From Cavanagh PR, Kram R. Mechanical and muscular factors affecting the efficiency of human movement. Med Sci Sports Exerc. 1985;17:326-331. Reprinted by permission of Williams & Wilkins. Figure based on data from Abbott BC, Bigland B, Ritchie JM. The physiological cost of negative work. J Physiol. 1952;117:380-390.) |
increases the mechanical stiffness of the joint and can provide considerable resistance to perturbating forces such as those that might be encountered while walking across a crowded room with a full glass. This same phenomenon may also play a role in protecting joints from traumatic forces.
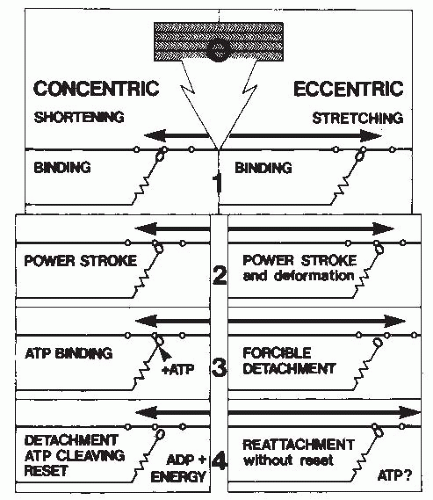 FIGURE 61-10. Schematic comparison of the crossbridge formation and cycling during concentric and eccentric work. ATP is required for the detachment and resetting of the crossbridges in concentric work but not in eccentric work. (From Cavanagh PR, Kram R. Mechanical and muscular factors affecting the efficiency of human movement. Med Sci Sports Exerc. 1985;17:326-331. Reprinted by permission of Williams & Wilkins. Figure based on data from White DCS. Muscle mechanics. In: Alexander RMcN, Goldspeak G, eds. Mechanics and Energetics of Animal Locomotion. London: Chapman and Hall; 1977:23-56.) |
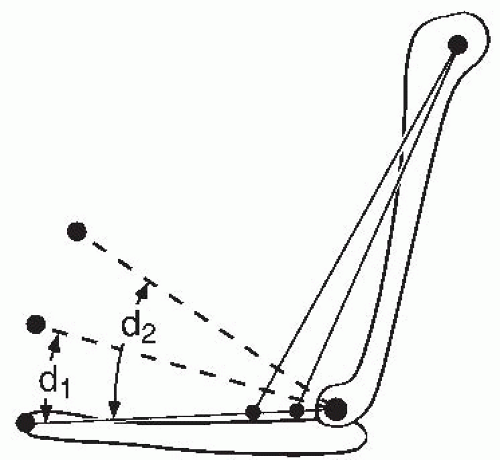 FIGURE 61-11. Schematic representation of the effect of tendon insertion site on excursion. The excursion from a more proximally inserted muscle may be double that of a more distally inserted muscle for the same amount of shortening. Whereas a tendon insertion closer to the center of rotation will allow greater excursion, torque production will be reduced for a given tension developed in the muscle. |
myosin filaments interact at the sarcomere level. Figure 61-13 shows the length-tension relationship for a single muscle fiber and the overlap of the actin and myosin filaments in a single sarcomere (40). As the overlap between actin and myosin increases, so does the tension production. Maximal tension is developed at lengths yielding maximal contact of actin and myosin filaments. As the sarcomere length decreases further, the actin filaments begin to overlap. It is believed that this interferes with crossbridge formation and causes a decline in tension development. Variations among sarcomeres and muscle fibers cause the length-tension curve to be more rounded for a whole muscle.
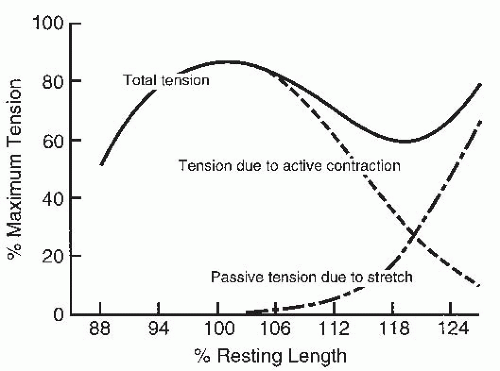 FIGURE 61-12. Length-tension diagram for passive stretch of an unstimulated muscle and total tension when the muscle is maximally stimulated. Active tension resulting solely from muscular contraction is obtained by subtracting the passive-stretch curve from the total-tension curve. Normal resting length is 100%. (Redrawn from Schottelius BA, Senay LC. Effect of stimulation-length sequence on shape of length-tension diagram. Am J Physiol. 1956;186:127-130.) |
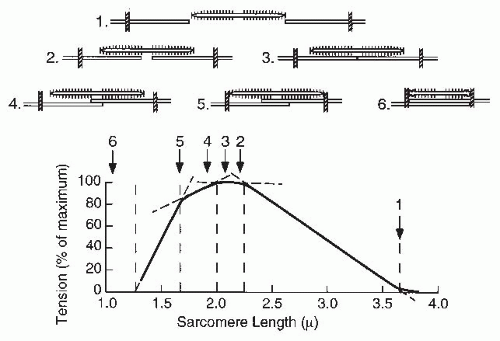 FIGURE 61-13. Relationship between sarcomere length and tension generation. The amount of overlap between actin and myosin filaments within each sarcomere is shown. The length of each sarcomere is given above, and the tension for each condition is shown below. Maximal tension is produced when there is the greatest overlap between filaments (points 2 and 3). Tension drops if the overlap is less or if the actin filaments contact each other. (Adapted from Gordon AM, Huxley AF, Julian FJ. The variation in isometric tension with sarcomere length in vertebrate muscle fibres. J Physiol. 1966;184:170-192.) |
movement commonly tested in the clinical setting (41). This figure demonstrates how maximal torque varies across movements when angular velocity is constant.
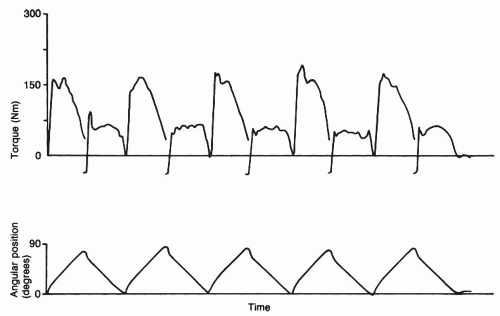 FIGURE 61-14. Real-time display of gravity-corrected torque and angular position during a knee extension-flexion isokinetic test. (From Baltzopoulos V, Brodie DA. Isokinetic dynamometry: applications and limitations. Sports Med. 1989;8:101-116.) |
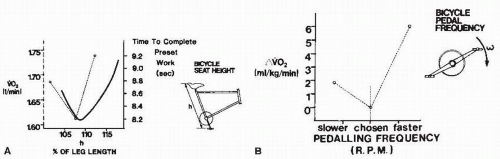 FIGURE 61-15. Optimal phenomenon in bicycling. Both seat height and pedaling frequency show an optimal point at which energy cost for producing a given power output is minimized. Maximal power output is also related to seat height by a parabolic curve. (From Cavanagh PR, Kram R. Mechanical and muscular factors affecting the efficiency of human movement. Med Sci Sports Exerc. 1985;17:326-331. Reprinted by permission of Williams & Wilkins. Figure based on data from Nordeen-Snyder K. The effect of bicycle seat height variation upon oxygen consumption and lower limb kinematics. Med Sci Sports. 1977;9:113-117; Hamley EJ, Thomas V. Physiological and postural factors in the calibration of the bicycle ergometer. J Physiol. 1967;191:55P-57P; Hagberg JM, Mullin JP, Giese MD, et al. Effect of pedaling rate on submaximal exercise responses of competitive cyclists. J Appl Physiol. 1981;51:447-451.) |
that optimizes the aerobic demands for walking a given distance (Fig. 61-17) (50).
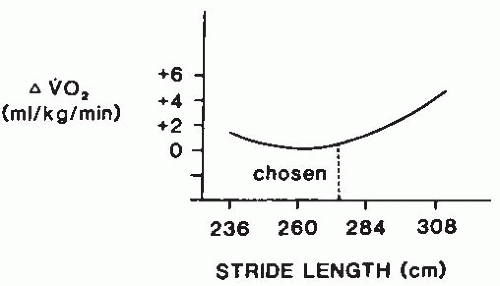 FIGURE 61-16. Optimal phenomena in running. For a given running speed, there is a stride length at which energy cost is minimized. The freely chosen stride length is typically close to optimal. (From Cavanagh PR, Williams KR. The effect of stride length variation on oxygen uptake during distance running. Med Sci Sports Exerc. 1982;14:30-35. Reprinted by permission of Williams & Wilkins.) |
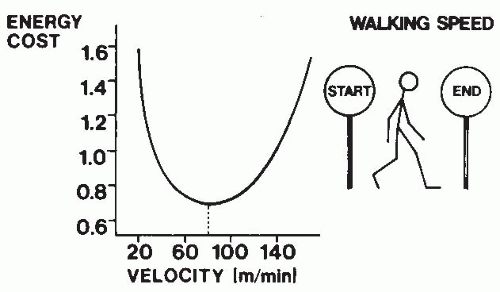 FIGURE 61-17. Optimal phenomena during walking. A speed exists at which energy cost per unit distance is minimized. Energy cost is shown here in units of kilocalories per kilogram body mass per meter traveled. (From Cavanagh PR, Kram R. Mechanical and muscular factors affecting the efficiency of human movement. Med Sci Sports Exerc. 1985;17:326-331. Reprinted by permission of Williams & Wilkins. Figure based on data of Ralston JH. Energy-speed relation and optimal speed during level walking. Int Z Angew Physiol. 1958;17:277-283.) |
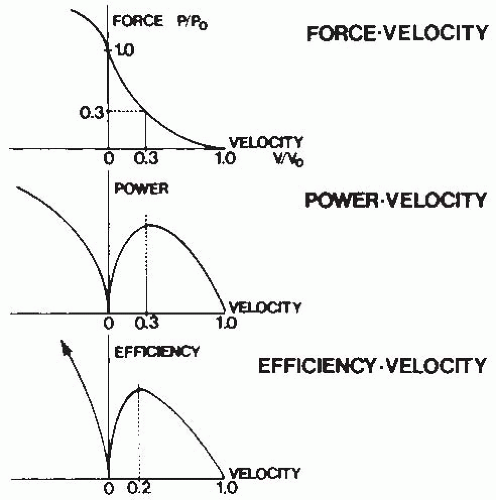 FIGURE 61-18. Optimal phenomena can be explained by derivations from the force-velocity relationship. The power-velocity curve is obtained from the product of force and velocity and demonstrates an optimal point. Energetic data allow generation of the efficiencyvelocity curve, which also shows an optimal point. (From Cavanagh PR, Kram R. Mechanical and muscular factors affecting the efficiency of human movement. Med Sci Sports Exerc. 1985;17:326-331. Reprinted by permission of Williams & Wilkins. Figure based on Hill AV. The maximal work and mechanical efficiency of human muscles and their most economical speed. J Physiol. 1922;56:19-41.) |
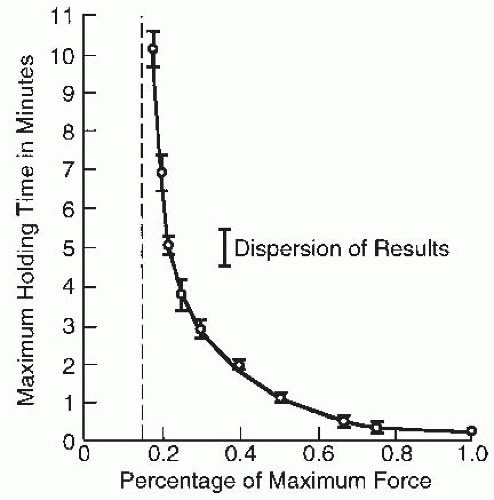 FIGURE 61-19. Isometric endurance as a function of percentage of maximal strength. (Adapted from Rohmert W. Ermittlung von erholungspausen fur statische arbeit des menschen. Int Z Angew Physiol. 1960;18:123-164.) |
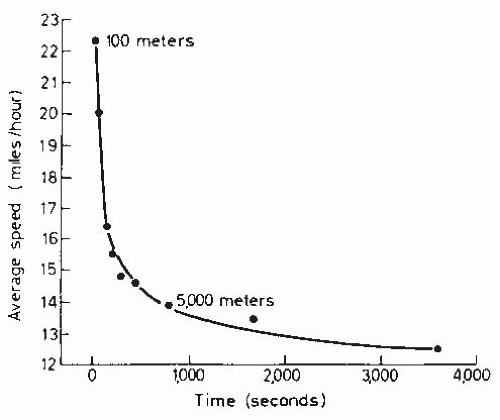 FIGURE 61-20. Relationship of running speed to exercise duration for world record runs. (From Simonson E. Recovery and fatigue. In: Simonson E, ed. Physiology of Work Capacity and Fatigue. Springfield, IL: Charles C Thomas; 1971:440-458. Reprinted by permission of Charles C Thomas, Publisher, Ltd.; Springfield, Illinois. Figure based on data from Lloyd BB. World running records as maximal performances: oxygen debt and other limiting factors. Circ Res. 1967;20-21(suppl 1):I218-I226.) |
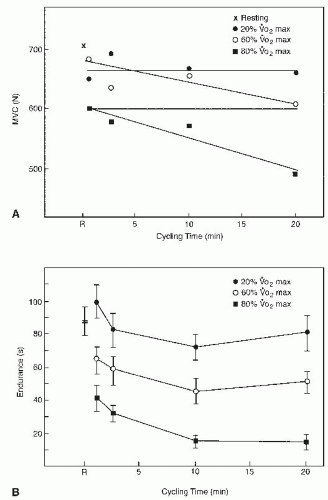 FIGURE 61-21. Effect of leg cycling at different intensities and durations on maximal isometric knee extension force generation (A) and isometric knee extension endurance at 40% of rested MVC (B). Mean isometric strength and endurance in the rested state are displayed at R. (With permission from Hoffman MD, Williams CA, Lind AR. Changes in isometric function following rhythmic exercise. Eur J Appl Physiol. 1985;54:177-183.) |
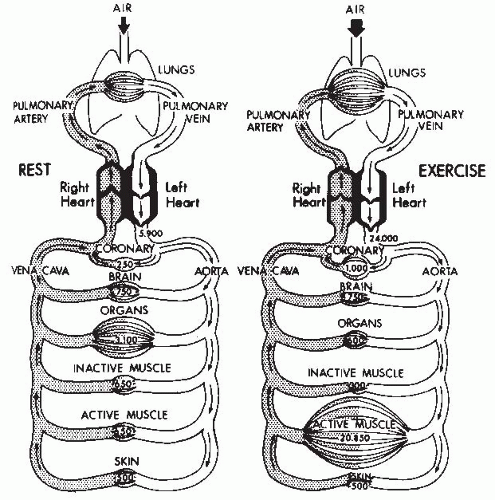 FIGURE 61-22. Schematic representation of the blood flow and distribution at rest and during maximal dynamic exercise. Exercise results in increases in blood flow to the exercising muscles and the coronary circulation but in reduced flow to the organs. Blood flow rates are indicated in milliliters per minute. (From Mitchell JH, Bloomqvist G. Maximal oxygen uptake. N Engl J Med. 1971;284:1018-1022. Reprinted by permission of The New England Journal of Medicine.) |
threefold increase in arteriovenous oxygen difference at maximal exercise (59).
of a volume load on the left ventricle (94,95). Although initial studies suggested that the magnitude of the presssor response to static exercise was primarily related to the percentage of MVC, most (but not all) subsequent studies suggest that the amount of muscle mass used also impacts positively on the pressor response.
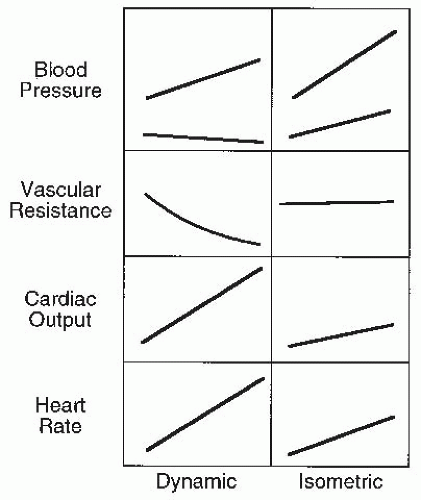 FIGURE 61-23. Schematic comparison of hemodynamic responses to dynamic and isometric exercise. (Adapted from Hanson P, Rueckert P. Hypertension. In: Pollock ML, Schmidt DH, eds. Heart Disease and Rehabilitation. 3rd ed. Champaign, IL: Human Kinetics; 1995: 343-356.) |
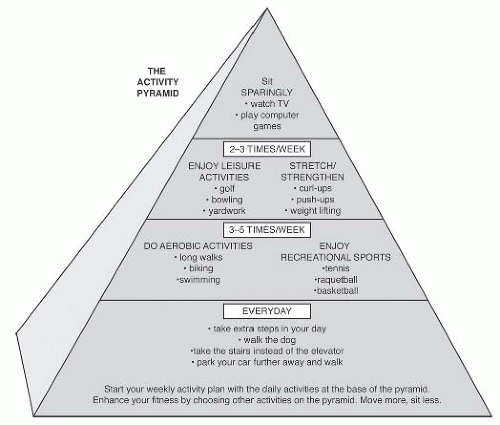 FIGURE 61-24. The Activity Pyramid is a model to facilitate education of the public about the adoption of a more active lifestyle. (Copyright 1997 Park Nicollet Health Source Institute for Research and Education. Reprinted by permission.) |
information on secondary prevention of cardiovascular disease, the reader is referred to the Chapter, “Cardiac Rehabilitation”.
(prediabetic state) reduced their incidence of DM during follow-up with a lifestyle diet/exercise intervention program designed to lose weight compared with those receiving normal care. This benefit was even more effective than that resulting from the administration of an oral antihyperglycemic agent.
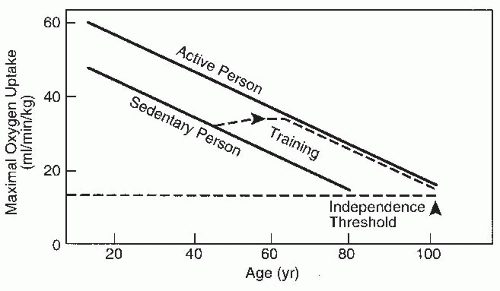 FIGURE 61-25. Demonstration of the effect of aerobic training on improving aerobic capacity and delaying its drop to a threshold where independent function can no longer be sustained. (Adapted from Shephard RJ. Exercise and aging: extending independence in older adults. Geriatrics. 1993;48:61-64.) |
coronary arterial vasoconstrictor response observed in atherosclerotic segments and thereby improve coronary artery blood flow. Aerobic conditioning has been recently reported to help prevent and restore age-related declines in endothelialdependent vasodilation in healthy people (185).
TABLE 61.4 Physiologic Adaptations to Aerobic Exercise Training as Observed in Resting and Exercise States | ||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
volume increases with training is not entirely clear but involves increased cardiac preload and probably enhanced myocardial contractility and relaxation (66,201). Aerobic training causes an increase in total blood volume that partially accounts for the increased cardiac preload. The extent of cardiac adaptations appears to be related to such training factors as the length, intensity, duration, and mode of training and the time of life at which training was initiated.
TABLE 61.5 Borg RPE 6-20 Scale | ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


