CHAPTER 5 Lumbar Musculature
Anatomy and Function
Physiology
The myofibrils attach to the sarcolemma at two ends, which connects one cell to its neighbor in the fiber structure. The myofibrils are highly organized, aligning longitudinally within the sarcolemma, which itself is indented by a motor axon at its myoneural junction. By convention, the smallest contractile subunit within the myofibrils is called the sarcomere. The sarcomere is composed of smaller subunits called myofilaments. The myofilaments are organized longitudinally with alternating light and dark striations (hence skeletal muscles are known as “striated muscle”) when visualized microscopically. The myofilaments within the sarcomere (smallest contractile subunit) are composed predominantly of two protein varieties: myosin and actin.1
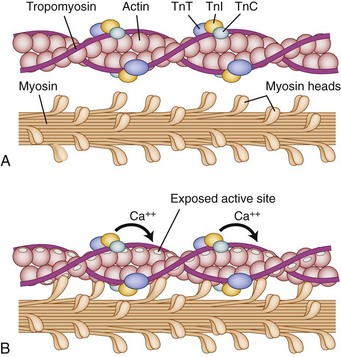
Under normal circumstances, contraction of striated muscle does not occur without a neural stimulus, whereas contraction of cardiac and most smooth muscle fibers can trigger adjacent fibers to contract without neural stimulation. The cellular mechanics of contractions are relatively simple: Actin filaments (occupying the light-colored I-band at rest) slide over the myosin filaments (found in the A-band and interdigitating with I-band at rest) until, with complete contraction, they completely overlap and eliminate the light H-band under microscopic visualization. The biochemical reactions are far more complex (Fig. 5–1). Contraction is initiated by release of acetylcholine at the myoneural junction, depolarizing the sarcolemma by changing its permeability to sodium and potassium ions. This sarcolemma stimulates release of calcium ions, sequestered in the sarcoplasmic reticulum, that bind to the troponin complex (C, T, and I). Calcium ions binding troponin induce a conformational change, which uncovers the binding portion of the actin filament. Myosin binds and unbinds actin, in concert with adenosine phosphate molecules (adenosine triphosphate [ATP] and adenosine diphosphate [ADP]), to induce the “racheting” of the myosin along the length of the actin filament.
A growing body of research shows that lactate is not a “dead-end metabolite,” and it is not the “mediator” of fatigue and inefficiency as widely published in the 1960s through the 1980s.2 On the contrary, current research implicates the hydrogen ion excess as the primary agent of diminished contractile power. The lactate ion may serve multiple roles in maintaining constant energy; recruiting new energy sources (gluconeogenesis); recruiting new vascularity (angiogenesis); and promoting a local cascade of healing, plasticity, and hyperplasia.2–4 A growing body of research based on the experimental work of Brooks,4 termed lactate shuttle theory, suggests that higher concentrations of lactic acid produced in the skeletal muscles stressed by exercise have significant increased benefit in remote tissue such as brain, liver, heart, peripheral nerves, and peripheral vasculature over baseline metabolism.2
The ATPase work of Engel5 in 1962 established a body of research showing the presence of distinctly different motor units within skeletal muscle. There are many myotype classification schemes based on histology, morphology, or function. In brief, the interaction between the type of myosin heavy chain (ATP binding site) and actin within individual sarcomeres is probably the greatest contributor to functional differences within myofibrils. The functional difference is related to the rate that the myosin heavy chains can repetitively bind ATP and release ADP under conditions of physiologic stress.6
Roughly divided, sarcomeres fall into one of three broad functional categories. Type I fibers have a slower “twitch” response with good fatigue resistance and lower tension development.7 Structurally, these groups of sarcomeres (known collectively as fibers) have rich capillary beds and high concentrations of mitochondrial enzymes with relatively low concentrations of glycogen and myosin ATPase. They seem ideally suited for aerobic activity with good fatigue resistance. These type I muscles predominate in areas that require aerobic or endurance demands. Type II muscle displays a fast twitch with good strength but relatively poor endurance compared with type I muscle fibers. Type II fibers can be subdivided further into type IIA, which still show a fast twitch response but a fatigue threshold between type I and type IIB, and type IIB, which show the fastest contractions, the highest tension development, and the most rapid onset of fatigue.7,8 Although other fiber subtypes continue to be identified, type I, type IIA, and type IIB show the major functional categories of voluntary skeletal muscles.7 Whether individual fibers have biochemical characteristics for high-intensity, short-duration contractile bursts or more sustained activity, each muscle is a heterogeneous, woven tapestry of all of the above-mentioned fiber subtypes. Any relative predominance of one particular fiber type is based on genetics, anatomic location of the muscle, demand on the muscle, age of the individual, nutrition, and multiple other external factors.9
Although the contribution to parental lineage to phenotypic expression may remain paramount in predicting functional potential, the actual fiber composition of muscle groups has great capacity for plasticity in response to environmental stress. Multiple studies show fiber conversion within major groups from type IIB to type IIA to be common.10 Other conversions, such as type I to either type IIA or type IIB, are less common and seen most often with denervation, immobilization, and profound deconditioning.8 There is emerging evidence of conversion of type II to type I in the electrical stimulation literature, but data remain sparse and confounded by the fact that type II muscle tends to atrophy with age. The relative absence of type II muscle may confound data trying to analyze conversion.11
A human infant is normally born with a full complement of muscle fibers that may continue to differentiate in childhood. Muscle fiber genesis seems to be biphasic. Myoblasts fuse to form fibrils at 8 to 10 weeks of gestation—primarily resulting in the formation of type I (slow-twitch) fibers. The second wave of fiber formation occurs at 15 to 18 weeks of gestation creating fast-twitch, type II fibers, which differentiate and change with early postpartum use.7 The growth and change in muscle size after adolescence seems to be due to increase in size of the fibers rather than increase in numbers (although controversial studies still contend that extreme muscle exertion may result in some new fiber development) that accompanies increased neuronal innervation and finer control of contractile force.
Although animal data suggest that hyperplasia and genesis of new muscle fibers may be possible with training and endogenous, hormonal feedback, these data have not been replicated in humans. Instead, increasing strength of contraction with training seems to be related not only to muscular factors, such as muscle size, fiber type, and fiber number, but also (and perhaps to a greater extent) to neural factors.12 The specific neural characteristics that change with training include frequency, extent, order, and synchrony of motor unit firing, with feedback to the spinal pyramidal tracts to control system efficiency—maximal force at minimal energy expenditure. Training, age, and certain anabolic hormones (either endogenous or exogenous) seem to exert the greatest effect on muscle plasticity—the characteristic rapid increase in strength and muscle diameter under the hormonal influence of puberty is a specific example.
Anatomy
The spine consists of a series of bilaterally symmetrical joints phylogenetically adapted for protection of the neural communications network linking brain to periphery.13 The critical role of the spine musculature in dynamically protecting and vitalizing these articulations with their passive ligamentous supports and accompanying neural transmission lines is infrequently acknowledged in biomechanical modeling. For this reason, a perspective based in evolutionary theory aids in understanding the complexity of lumbar spine musculature.14 From an evolutionary standpoint, it seems that all land-dwelling vertebrates (including mammals) evolved from ocean-dwelling cousins; in the ocean, gravitational force acted differently when contributing to function and form. Large paravertebral spine muscles likely developed to provide lateral (coronal plane) flexion-extension to propel bodies through water, as shown by the lateral tail motion of the fish. This form of locomotion was initially preserved in amphibians and land reptiles (even after the development of extremities), as seen in the Crocodylia species including the modern alligator and crocodile. Reptiles with legs (currently living) all propel themselves through lateral spinal motion in which propulsion is achieved by contracting spine muscles to create alternating coronal convexities on one side of the spine that allow the ipsilateral foreleg to move forward while the contralateral hindfoot (on the concave side) is brought closer to the contralateral foreleg in preparation for the next reciprocal lateral movement that repeats the motion-event.
Mammalian adaptations resulted in a 90-degree evolutionary shift to the sagittal spinal motion characteristic of all four-legged mammals (the platypus and echidna being partial exceptions), which presumably provides an advantage during land locomotion, allowing for explosive growth of the class Mammalia. Land mammals that subsequently retreated to the sea (e.g., whales, porpoises, seals) maintain the sagittal orientation of spinal locomotion resulting in the 90-degree rotation of the tail fluke (when compared with fish), even after adaptive pressure resulted in other changes to their extremities. The later adaptation of humans to full bipedal stance and locomotion presumably necessitated a lordotic lumbar spine, kyphotic thoracic spine, and lordotic cervical spine for balance and ambulation (Fig. 5–2).
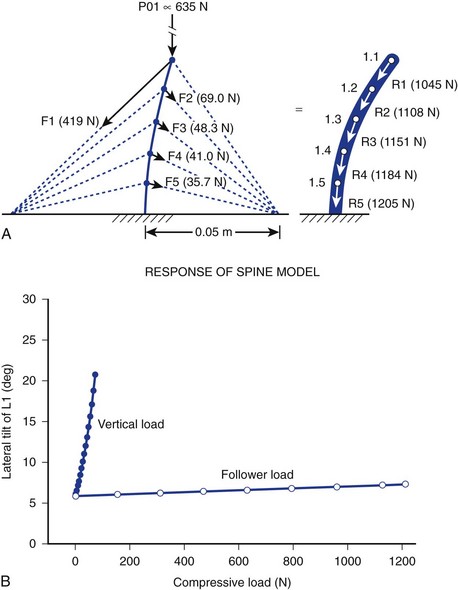
It is further theorized that the lordotic curve converts lateral bending to produce torsion at the hips, adding propulsion efficiency to the balanced human gait that would be impossible without lumbar lordosis. Ambulation without lumbar lordosis leads to the shuffling strides of the upright apes, whose gait is clearly dissimilar to that of humans. Laboratory modeling strongly suggests that cocontraction of spinal and abdominal muscles is the primary generator of the curvilinear structure of the spine that sustains logarithmically more force than “straight” models of spine motion (>1200 N vs. 100 N). The theory holds that instantaneous, axial-rotational forces between segments in “straight-spine” models lead to rapid failure when the spine is progressively loaded.15
This model focuses on the dynamic contribution of spine muscles to create a compressive-stabilizing force through bony and ligamentous structures that withstands physiologic forces modeled in sagittal motion. Similar to taut “guy wires” or tensioned cables adding structural stability to allow gauzy tent material to withstand 90 mph winds, muscle tension is hypothesized by the “follower-load” theory to provide a stabilizing force (in at least one plane of motion—sagittal) that directs the force vector to pure compression of the motion segment, minimizing shear (shear force being implicated in degenerative cascade).16
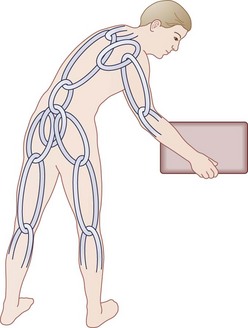
Another unique human evolutionary adaptation for manipulation of objects near the ground from a bipedal stance involves the ability to bend efficiently from the waist in combination with squatting and hunkering. A stable “biomechanical chain,” transferring force efficiently from hands through arms, shoulder girdle, spine, pelvis, legs, and feet to make ground contact, is necessary (Fig. 5–3). In this functional concept, the muscle acts as a dynamic stabilizer of the biomechanical chain in several ways. During flexion-extension of the lumbar spine, the co-tensioning abdominal muscles at a distance (force plus lever arm) allows maintenance of a “balance point” at each individual motion segment. Next, coupled anterior and posterior forces have a net compressive (downward) force, which balances motion at the instantaneous axis of rotation for each motion segment—maintaining compression at the disc and minimizing angular change. The net muscle tension force downward serves to offset any other forces, maintaining the force vector perpendicular to the disc’s plane similar to the tent “guy-wires” explained previously.16,17 Muscles supply dynamic and static downward force to create a form that allows maximal load bearing (for bipedal carrying and lifting) capacity, while maintaining function that allows maximal efficiency of muscle energy output—maintaining a plumb line (not working against gravity to maintain posture), sharing tension bands to distribute loads, coupling forces when motion is required, and distributing load/work among the other osteoligamentous static structures.
These observations lead to multiple functional inferences, one of which is the tendency of the spine to “hang on its ligaments” in an efficient, muscle-sparing manner frequently observed in “stooped laborers.” Efficiency of motion is documented by the way most normal subjects make the universal unconscious choice of allowing spine flexion to precede hip flexion when bending forward at the waist, eventually resulting in an EMG “silent period” when the spine is fully flexed and stabilized by posterior ligaments.14,18,19
The small interconnecting vertebrae motion segments with the multiplanar motions of the three-joint complex produce difficulties when assigning specific uniaxial functions to individual groups of muscles. The erector spinae group of muscles are generally thought of as extensors of the spine. Functioning unilaterally, they may also be powerful abductors or lateral stabilizers (assisting with locomotion) and have been shown to function to some extent in spine derotation.18 Similarly, lateral abdominal musculature (internal and external oblique and transversus abdominis) may act as spine flexors and extensors (working through the lumbosacral fascia).14 The “girdling” abdominal muscles are also powerful spine rotators and assist in abduction and lateral stabilization.20 As noted previously in this chapter, the force-coupling of spinal, pelvic, and abdominal muscles acting in concert increases compressive load through the disc and vertebral bodies to minimize shear force and imbalance axially across the three-joint complex.
In describing the gross anatomy of the spinal muscular functional unit, the extreme importance of the posterior stabilizing structures cannot be overemphasized in spinal functional integrity. The intrinsic lumbar musculature is only a part of the functional unit. The lumbodorsal fascia, interspinous ligaments, and facet joint capsules are crucial structures providing constraint and fulcrum points.21 There is growing appreciation among some surgeons to maintain the integrity of these collagenous and elastic structures, which may decrease adjacent segment shear, dysfunction, or instability.
Mammals as a class share many characteristics, including those of the spine musculature, that may account for the evolutionary success of the class. The most superficial layers of spine musculature, extensions of the functional unit of the shoulder girdle, include muscles such as the serratus posterior and latissimus dorsi. In contrast to other spinal musculature, the modified proximal function of these muscles is matched by innervation from the proximal spinal cord. The true lumbar spinal muscles, by contrast, have segmental innervations that arise from the posterior rami of the contiguous spinal nerves (the same nerves that perceive proprioceptive input from facet capsules, posterior ligaments, and peripheral anulus). Although true spinal muscles function together, their most characteristic differentiating factor is span length. The deepest muscles, such as the interspinalis, span only a single segment. The most superficial muscles may traverse a large portion of the entire spinal column. Controlled, coordinated action of individual vertebrae is a critical part of spine function, whereas loss of appropriate musculoligamentous control may contribute to various pathologic syndromes, such as segmental instability, segmental rigidity, facet syndromes, and perhaps even discogenic pain.22
Musculature of the Lumbar Spinal Functional Unit
Intrinsic Muscles
Erector Spinae
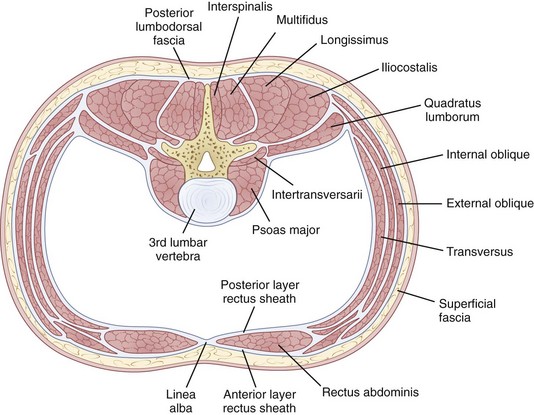
The erector spinae is a large and superficial muscle that lies just deep to the lumbodorsal fascia and arises from an aponeurosis on the sacrum, iliac crest, and thoracolumbar spinous processes.13 The muscle mass is poorly differentiated, but divides into three sections in the upper lumbar area: (1) The iliocostalis is most lateral and inserts into the angles of the rib; (2) the longissimus, the intermediate column, inserts into the tips of the spinous processes of thoracic and cervical vertebrae; (3) the spinalis is most medial and inserts into spinous processes of the cervical and thoracic vertebrae (Fig. 5–4). The innervation of all the lumbar paraspinal muscles is from the dorsal rami of the nerve root as it exits at the most approximate level and overlaps proximally and distally along the muscle length.
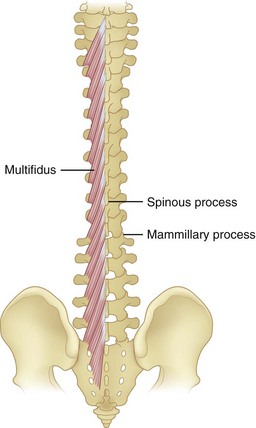
Multifidi
Multifidi are a series of small muscles, best developed in the lumbar spine, that originate on the mammillary processes of the superior facets and run upward and medially for two to four segments, inserting on the spinous processes (Fig. 5–5). This orientation produces greater capacity for rotation and abduction, in addition to extension. They also share a multilevel innervation (similar to the erector spinae bundle or group) where function is maintained even with injury to the proximate dorsal rami.
Quadratus Lumborum
The quadratus lumborum is the most lateral of the lumbar muscles (see Fig. 5–4); it originates on the iliac crest and iliolumbar ligament and runs obliquely to insert into the lowest rib and transverse processes of the upper four lumbar vertebrae. Its orientation provides strong abduction motor and stabilizer properties. The innervation is from the ventral rami of T12-L1-L2-L3 roots.
Psoas and Iliacus Muscles
The psoas major, although usually thought of primarily as a hip flexor, has a direct effect on the vertebral column because it originates bilaterally from the vertebral bodies and posterior aspects of the transverse processes, providing the only intrinsic spinal muscle acting anterior to the sagittal axis. Yet, paradoxically, the psoas is usually an intersegmental extensor in the mid-lumbar spine, even as it flexes at the lumbosacral junction in the process of increasing the lumbar lordosis. It is an important spine stabilizer in sitting and standing.23 Acting asymmetrically, the psoas may produce abduction or abduction-resistance to maintain coronal balance. Pathophysiologically, low back or pelvic pain may result from contracture or spasm of the iliopsoas producing combined hip and lumbosacral junction flexion. As in other situations, appropriately targeted stretches, followed by strengthening, may produce dramatic improvement.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


