CHAPTER 2 Gross Anatomy of the Shoulder
HISTORY
During the Renaissance, a unique style of painting called glazing was developed. The artist applied to the canvas thin, transparent coats of different colors. The deep layers of paint, viewed through the more superficial layers, yielded a color variant and perception of depth that was not attainable with a single layer of paint. El Greco, Titian, and Rubens were among the artists who used glazing.
Before the Renaissance, the main barriers to anatomic study were religious and personal proscriptions against the dissection of human cadavers and different philosophic ideas regarding the laws of nature. As a result of these prejudices, early contributions were not ad-vanced, and because they were not reconfirmed, they were lost to humanity. In a dialogue written in the warring states period (5th century bc), Huang Ti described the unidirectional flow of blood in arteries and veins. Huang Ti, the Yellow Emperor of China who lived around 2600 bc, is the mythologic father of Chinese medicine.1–3
This and other indications of anatomic study were buried in a heavily philosophic treatise about the yin and yang influences on the human body. (Recent work in traditional Chinese medicine correlates the yang acupuncture meridians with the course of postaxial nerve branches of the posterior cord and the yin meridians with the preaxial nerve branches of the medial and lateral cords.4) Over the centuries, this Daoist orientation led to a de-emphasis on surgical anatomy and a resultant prejudice against surgery. Although Chinese anatomic observations were subsequently carried out by physicians who witnessed executions, the “death of a thousand cuts,” and bodies disinterred by dogs after an epidemic, the philosophic climate was unfavorable for propagation and use of this information.1 Therefore, a lasting description of circulation awaited rediscovery by John Harvey in the more receptive atmosphere of the 17th century, approximately 2000 years later.
The handling of dead bodies was prohibited in India, except when preparing them for cremation; however, Sustruta, in the 6th century bc, devised a frame through which flesh was dissected away from the deeper layers with stiff brooms.5,6 He correctly described the two shoulder bones (clavicle and scapula) at a time when the West thought of the acromion as a separate bone. In the same era, Atroya fully described the bones of humans. Alcmaeon performed animal dissection around 500 bc in Greece.6 The significance of these discoveries was lost because further study was not conducted.
Hippocrates was probably the first physician whose ideas regarding shoulder anatomy were perpetuated.7 Writing in the 5th century bc, his discussion of articulations began with the shoulder, and much of his work focused on this joint. His writings applied to clinical rather than basic science. Although Hippocrates referred to “the unpleasant if not cruel task” of dissecting cadavers, we must assume that he witnessed dissections because he gave explicit instructions for obtaining exposure of portions of the shoulder anatomy to prove a clinical point.6,8 He also described the position of nerves of the axilla when discussing his burning technique for treatment of anterior dislocation of the shoulder, and in his assessment of patients, he noted that some have a “predisposing constitution” to dislocation. He demonstrated knowledge of acromioclavicular separation, palsies of the shoulder, and growth plate fractures.8
Herophilus (circa 300 bc), the father of anatomy, dissected some 600 cadavers at the medical school in Alexandria and started an osteology collection. This collection is the first recorded evidence of what might be called a scientific approach in that he dissected more than just a few cadavers and performed his dissections for description rather than for pathologic analysis.6 Early authors such as Celsus accused him of vivisection, but this allegation is unjustified.6 His permanent anatomic collections, particularly of bones, contributed to the medical knowledge of the late Greek and early Roman eras. In the late Republic and early Empire, new proscriptions against dissection, some of which were written into law, stemmed the progress that had been made. Celsus (30 bc-41 ad), who was not a physician but an encyclopedist, collected the medical knowledge of the day and advocated the performance of human dissection, but the prohibition continued.3,6
Despite such barriers to the study of human anatomy, advances were made sporadically by gifted physicians in unique situations. Galen, a Greco-Roman physician who practiced in Pergamum and then in Rome during the 2nd century ad, contributed greatly to the early knowledge of anatomy. He is the father of the study of clinical anatomy and, because he was the surgeon for gladiators in Pergamum, might also be considered the father of sports medicine.9 His writings on the “usefulness of the human body parts” contain the earliest effort at detailed anatomy of the shoulder. In discussing the bones and joints of the shoulder, he described the thin ligaments of the glenohumeral joint and observed that the rim of the glenoid was often broken with dislocation. Galen attributed the frequency of dislocation of the shoulder, in comparison to other joints, to an “antagonism between diversity of movement and safety of construction.” He described the anatomic principle of placing joints in a series to increase motion so that “the additional articulation might supply the deficiency of the first articulation.” He also provided a complete description of all the muscles and their subdivisions, although he did not apply the Latin names used today.9
In describing the nerves, Galen referred to a sympathetic trunk and a plexus but did not apply names or recognize a standard construction of the brachial plexus. Instead, he considered the plexus to be a necessary method of strengthening support to the nerves. He described terminal branches of the brachial plexus, including the dorsal scapular, axillary, median, ulnar, and radial nerves. Galen also described the accessory spinal nerve. He noted the axillary artery, carotid artery, and lymph glands about the shoulder.9 In short, he provided an impressive outline of shoulder anatomy, even by 21st-century standards.
Many differences between Galen’s writings and modern descriptions relate to the fact that he performed only animal dissection. He perpetuated the erroneous idea that the acromion is a separate bone, a concept that continued into the Renaissance. In addition, his writings on the acromion probably do not refer to the acromion as we know it but to the acromioclavicular joint.9
Shortly after Galen’s time, Christianity became dominant in the Roman empire, which led to intensification of the preexisting laws against cadaveric dissection.3 For centuries, no anatomic studies were performed on human material in Europe or in the Muslim empire. Although the Muslims were more successful in preserving the Galenic writings, their religion prohibited illustration. Perhaps the very completeness of Galen’s studies contributed to the suppression of anatomic studies. Centuries passed with no new knowledge of human anatomy being acquired.6
When the Greek and Roman literature was reintroduced from the East, scholasticism was the dominant academic philosophy in the West. The scholastic philosophers Abelard and Thomas Aquinas depended heavily on deductive reasoning for any original contributions that they made and did not use observation or experimentation.3,6 In view of the rich sources that reappeared from the East, one can understand how a scholar could absorb a much larger volume of information from ancient writings than from the slow and laborious process of experimentation.
Finally, during the Black Plague of 1348, the papacy allowed necropsy to be performed for the first time to elucidate the cause of death from the plague but not for anatomic study. Interestingly, when dissection for the purpose of teaching anatomy was reintroduced, it was not investigational but simply demonstrated the precepts of Galen.6
One of the greatest leaps in anatomic illustration occurred among Renaissance painters interested in accurate representation, some of whom dissected in secret and became serious students of anatomy (Fig. 2-1). Although their purpose was illustration and not new discovery, the increase in accuracy of illustrations in the notebooks of Leonardo da Vinci in comparison to older anatomic textbooks is quite remarkable.6 Leonardo, in his early notebooks, was seeking to illustrate ideas from Mondino’s dissection manual, which Mondino derived from Galen. The bulk of his drawing, however, is made up of his own observations, often independent of Galen.10 Leonardo also recognized the value of dissecting multiple specimens:
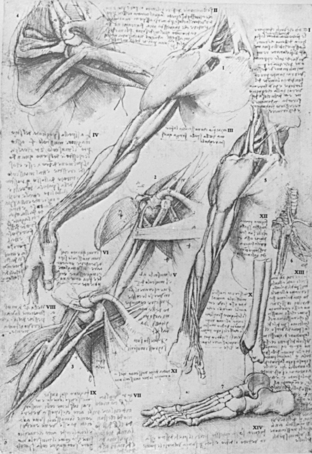
And you who say that it is better to look at an anatomical demonstration than to see these drawings, you might be right, if it were possible to observe all the details shown in these drawings in a single figure, in which with all your ability you will not see … nor acquire a knowledge … while in order to obtain an exact and complete knowledge of these … I have dissected more than ten human bodies.10
In his shoulder illustrations, he shows fused and unfused acromia without notation regarding which was abnormal. His notations are largely instructions to himself, such as “draw the shoulder then the acromion” rather than observations of the incidence of variations.10
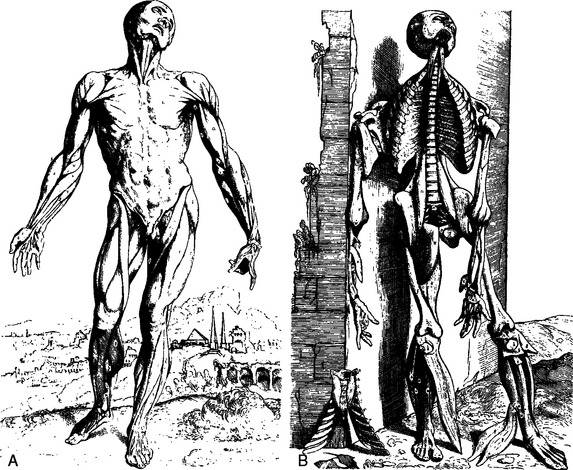
Anatomic study with clear illustration steadily increased over the next century. In 1537, Pope Clement VII endorsed the teaching of anatomy, and in 1543, Vesalius published his textbook De Fabrica Corporis Humani (Fig. 2-2).11 Vesalius, although criticized for questioning the work of Galen during his early teaching career, was able to correct some of Galen’s misconceptions. He described the ge-ometry of muscles and contributed the concept of the dynamic force of the body, as illustrated in a vivid portrayal of progressive muscle dissection of the cadaver. In his artist’s illustrations, the cadaver appears to lose more tone as each muscle layer is removed until, finally, in the last picture, the cadaver has collapsed against a wall. Vesalius demonstrated the vessels of the shoulder, and his drawings include an accurate illustration of the brachial plexus. The only element missing is the posterior division of the lower trunk (Fig. 2-3). He accurately portrayed rotation of the fibers of the costal portion of the pectoralis major.11 His drawings also include material from comparative anatomy and indicate where these structures would lie if they were present in a human. This work was the starting point of scientific anatomy.6
The functions of muscles were deduced early from their shortening action and their geometry. While caring for patients during an anthrax epidemic, Galen asked them to perform certain arm motions. Because of his knowledge of anatomy, he was able to determine, without painful probing, exactly which muscle he was observing at the base of the anthrax ulcers typical of the disease and how close these ulcers were to vital nerves or vessels.6 Modern biomechanics still benefits from study of the geometry of muscles.12
The dynamic study of muscles was made possible with the development of electrical equipment. DuBois-Reymond invented the first usable instrument for the electrical study of nerves and muscles in the early 19th century, and Von Helmholtz first measured the speed of nerve conduction.6 Duchenne studied the action of muscles by electrically stimulating individual muscles through the skin, and like his predecessors, he began with the shoulder and emphasized that joint (Fig. 2-4). Duchenne also recognized that muscles rarely act individually and that this limited the accuracy of his method. He studied all the superficial muscles of the shoulder, including the trapezius, rhomboid, levator scapulae, serratus anterior, deltoid, supraspinatus, infraspinatus, teres minor, subscapularis, latissimus dorsi, pectoralis major, teres major, and triceps.6,13,14 Subsequent developments in electromyography enabled researchers to measure muscle activity initiated by the patient.14
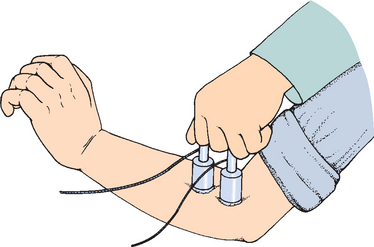
(From Duchenne GB: Physiology of Movement, ed and trans EB Kaplan. Philadelphia: WB Saunders, 1959.)
Functional anatomy was further elucidated by the science of physics. Aristotle studied levers and geometry and wrote about the motion of animals. Galen wrote about muscle antagonists. Leonardo da Vinci discussed the concept of centers of gravity in his notebooks. However, it was Sir Isaac Newton’s physics that made possible the studies that we perform today. In the late 19th century, Eadweard Muybridge15 published photographic studies of a horse in motion, and he later photographed human motion in rapid sequence to examine the action of the various levers of the body (Fig. 2-5). It was rapid-sequence photography that elucidated the synchrony of glenohumeral and scapulothoracic motion.6,16 Braune and Fischer first applied Newtonian physics to functional anatomy.17,18 In their classic studies, they used cadavers to establish the center of gravity for the entire body and for each segment of the body, the first detailed study of the physics of human motion based on such information.7 Awareness of the great motion of the shoulder made it one of the first areas where overuse was described.19

The first thorough study of any joint combining all the techniques of the historical investigators was performed on the shoulder by Inman, Saunders, and Abbott.20 This landmark work used comparative anatomy, human dissection, the laws of mechanics, photography, and the electromyogram.20 All subsequent publications on the function of the shoulder might support or contradict findings in this study, but all cite it.12
Cadaveric dissection and other research continue to add to our knowledge of the shoulder and to our understanding of the findings of these early giants in the field. Exciting studies are presented at meetings and wherever the shoulder is a topic of discussion. These studies are stimulated by the same three sources that have inspired anatomists throughout history. The first stimulus is a new disease or a new understanding of an old disease. The premiere example here is the studies of impingement syndrome. The second stimulus is the invention of new technology for treatment, such as when the arthroscope activated renewed interest in variations in the labrum and ligaments of the shoulder.21 The third is the invention of a new technique to study anatomy. In the past, this has occurred with Duchenne’s electrical stimulus, the electromyogram, and the fluoroscope.16 More recently, biochemistry, the electron microscope, magnetic resonance imaging (MRI), and laser Doppler vascular evaluation have stimulated new interest. All these techniques have deepened our understanding of previous findings in the shoulder rather than totally altered the picture (Fig. 2-6).

The final layer of paint in our portrait of shoulder anatomy has not been applied, and we are unlikely to see the day when further study is not required. Scholasticism appears to be creeping into medical school education. This trend is probably an unfortunate byproduct of the information surge of the late 20th and early 21st century, a phenomenon similar to the reappearance of Galen’s writing in the era of scholasticism. More time is spent memorizing accumulated information at the cost of research experience. Although at present a revolution in surgical technique and technology has sent a generation of surgeons back to the anatomy laboratory,21 one must not forget to encourage an interest in anatomy research in medical students.
BONES AND JOINTS
The orthopaedic surgeon thinks of bones primarily as rigid links that are moved, secondarily as points of attachment for ligaments, muscles, and tendons, and finally, as the base on which to maintain important relation-ships with surrounding soft tissue. Treatment of fractures has been called the treatment of soft tissues surrounding them.22 In relation to pathology, bones are three-dimensional objects of anatomy that must be maintained or restored for joint alignment. Bones exist in a positive sense to protect soft tissue from trauma and provide a framework for muscle activity. In a negative sense, they can act as barriers to dissection for a surgeon trying to reach and repair a certain area of soft tissue. Loss of position of the bone can endanger soft tissue in the acute sense, and loss of alignment of the bone can endanger the longevity of the adjacent joints.
This great range of motion is distributed to three diarthrodial joints: the glenohumeral, the acromioclavicular, and the sternoclavicular. The last two joints, in combination with the fascial spaces between the scapula and the chest, are known collectively as the scapulothoracic articulation.20 Because of the lack of congruence in two diarthrodial joints (the acromioclavicular and sternoclavicular joints), motion of the scapulothoracic articulation is determined mainly by the opposing surfaces of the thorax and scapula. About one third of the total elevation takes place in this part of the shoulder; the remainder occurs in the glenohumeral cavity. The three diarthrodial joints are constructed with little bony stability and rely mainly on their ligaments and, at the glenohumeral joint, on adjacent muscle. The large contribution of the scapulothoracic joint to shoulder function and to axial body mechanics has been emphasized since the 1990s.
The division of motion over these articulations has two advantages. First, it allows the muscles crossing each of these articulations to operate in the optimal portion of their length-tension curve. Second, the glenohumeral rhythm allows the glenoid to be brought underneath the humerus to bear some of the weight of the upper limb, which decreases demand on the shoulder muscles to suspend the arm. Such division of motion is especially important when the muscles are operating near maximal abduction and they are at that point in their length-tension curve where they produce less force.23,24 Study of the ultrastructure of ligaments and tendons about the shoulder is in its infancy, but preliminary studies show little difference in terms of collagen biochemistry and fiber structure.25,26
Sternoclavicular Joint
The sternoclavicular joint, which is composed of the upper end of the sternum and the proximal end of the clavicle, is the only skeletal articulation between the upper limb and the axial skeleton.27 In both the vertical and anteroposterior dimensions, this portion of the clavicle is larger than the opposing sternum and extends superiorly and posteriorly relative to the sternum.27,28 The prominence of the clavicle superiorly helps create the suprasternal fossa. The sternoclavicular joint has relatively little bony stability, and the bony surfaces are somewhat flat. The ligamentous structures provide the stability of the joint. The proximal surface of the clavicle is convex in the coronal plane but somewhat concave in the transverse plane. The joint angles posteromedially in the axial plane. In the coronal plane, the joint surface is angled medially toward the superior end; the joint surfaces are covered with hyaline cartilage. In 97% of cadavers, a complete disk is found to separate the joint into two compartments (Fig. 2-7). The disk is rarely perforated.29,30 The intra-articular disk is attached to the first rib below and to the superior surface of the clavicle through the interclavicular ligament superiorly. The disk rarely tears or dislocates by itself.31
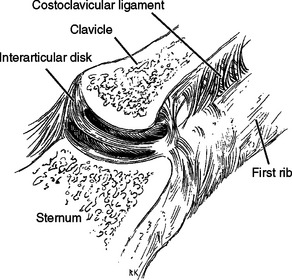
The major ligaments in the joint are the anterior and posterior sternoclavicular or capsular ligaments (Fig. 2-8). The fibers run superiorly from their attachment to the sternum to their superior attachment on the clavicle. The most important ligament of this group, the posterior sternoclavicular ligament, is the strongest stabilizer to the inferior depression of the lateral end of the clavicle.32 The paired sternoclavicular ligaments are primary restraints so that minimal rotation occurs during depression of the clavicle.
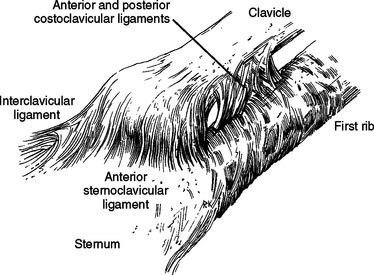
The interclavicular ligament runs from clavicle to clavicle with attachment to the sternum, and it may be absent or nonpalpable in up to 22% of the population.25 The ligament tightens as the lateral end of the clavicle is depressed, thereby contributing to joint stability.
The anterior and posterior costoclavicular ligaments attach from the first rib to the inferior surface of the clavicle. The anterior costoclavicular ligament resists lateral displacement of the clavicle on the thoracic cage, and the posterior ligament prevents medial displacement of the clavicle relative to the thoracic cage.33 Cave thought that these ligaments acted as a pivot around which much of the sternoclavicular motion takes place.34 Bearn found that they were not the fulcrum in depression until after the sternoclavicular ligaments were cut. They are the principal limiting factor in passive elevation of the clavicle and are a limitation on protraction and retraction.32 Perhaps the costoclavicular ligaments allow the good results reported for proximal clavicle resection.33
In the classic study on stability of the sternoclavicular joint, Bearn32 found that the posterior sternoclavicular or capsular ligament contributed most to resisting depression of the lateral end of the clavicle. He performed serial ligament releases on cadaver specimens and made careful observations on the mode of failure and the shifting of fulcrums. This qualitative observation is a useful addition to computerized assessment of joint stability.
Although reliable electromyographic studies demonstrate that the contribution of the upward rotators of the scapula is minimal in standing posture, permanent trapezius paralysis often leads to eventual depression of the lateral end of the scapula relative to the other side, although this depression may be only a centimeter or two.32 Bearn’s experiment probably should be replicated with more sophisticated equipment to produce length-tension curves and to quantitatively test the response of the joint to rotational and translational loading in the transverse and vertical axes, as well as the anteroposterior axis that Bearn tested qualitatively.32
Motion occurs in both sections of the sternoclavicular joint: elevation and depression occur in the joint between the clavicle and the disk,27 and anteroposterior motion and rotatory motion occur between the disk and the sternum. The range of motion in living specimens20 is approximately 30 to 35 degrees of upward elevation. Movement in the anteroposterior direction is approximately 35 degrees, and rotation about the long axis is 44 to 50 degrees. Most sternoclavicular elevation occurs between 30 and 90 degrees of arm elevation.20 Rotation occurs after 70 to 80 degrees of elevation. Estimation of the limitation of range of motion as a result of fusion is misleading because of secondary effects on the length- tension curve of the muscles of the glenohumeral joint and the ability of the glenoid to help support the weight of the arm. Fusion of the sternoclavicular joint limits abduction to 90 degrees.12,16
The blood supply to the sternoclavicular joint is derived from the clavicular branch of the thoracoacromial artery, with additional contributions from the internal mammary and the suprascapular arteries.27 The nerve supply to the joint arises from the nerve to the subclavius, with some contribution from the medial supraclavicular nerve.
Immediate relationships of the joint are the origins of the sternocleidomastoid in front and the sternohyoid and sternothyroid muscles behind the joint. Of prime importance, however, are the great vessels and the trachea (Fig. 2-9), which are endangered during posterior dislocation of the clavicle from the sternum’a rare event that can precipitate a surgical emergency.22,35,36
An open epiphysis is a structure not commonly found in adults. The epiphysis of the clavicle, however, does not ossify until the late teens and might not fuse to the remainder of the bone in men until the age of 25 years.32,36 Therefore, the clavicular epiphysis is a relatively normal structure within the age group at greatest risk for major trauma. The epiphysis is very thin and not prominent, which makes differentiation of physeal fractures from dislocations difficult. Instability of the sternoclavicular joint can result from trauma but, in some persons, develops secondary to constitutional laxity.37
Clavicle
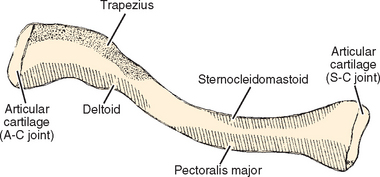
The clavicle is a relatively straight bone when viewed anteriorly, whereas in the transverse plane, it resembles an italic S (Fig. 2-10).38 The greater radius of curvature occurs at its medial curve, which is convex anteriorly; the smaller lateral curve is convex posteriorly. The bone is somewhat rounded in its midsection and medially and relatively flat laterally. DePalma38 described an inverse relationship between the degree of downward facing of the lateral portion of the clavicle and the radius of curvature of the lateral curve of the clavicle.
The obvious processes of the bone include the lateral and medial articular surfaces. The medial end of the bone has a 30% incidence of a rhomboid fossa on its inferior surface where the costoclavicular ligaments insert and a 2.5% incidence of actual articular surface facing inferiorly toward the first rib. The middle portion of the clavicle contains the subclavian groove where the subclavius muscle has a fleshy insertion (Fig. 2-11). The lateral portion of the clavicle has the coracoclavicular process when present.
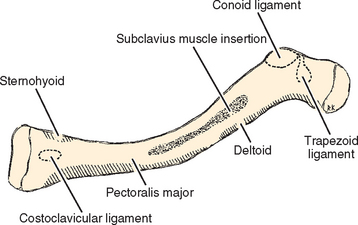
The clavicle has three bony impressions for attachment of ligaments. On the medial side is an impression for the costoclavicular ligaments, which at times is a rhomboid fossa. At the lateral end of the bone is the conoid tubercle, on the posterior portion of the lateral curve of the clavicle and the trapezoid line, which lies in an anteroposterior direction just lateral to the conoid tubercle. The conoid ligament attaches to the clavicle at the conoid tubercle and the trapezoid ligament attaches at the trapezoid line. The relative position of these ligament insertions is important in their function.27,28,38
Muscles that insert on the clavicle are the trapezius on the posterosuperior surface of the distal end and the subclavius muscle, which has a fleshy insertion on the inferior surface of the middle third of the clavicle. Four muscles originate from the clavicle: The deltoid originates on the anterior portion of the inner surface of the lateral curve; the pectoralis major originates from the anterior portion of the medial two thirds; the sternocleidomastoid has a large origin on the posterior portion of the middle third; and the sternohyoid, contrary to its name, does have a small origin on the clavicle, just medial to the origin of the sternocleidomastoid.
Functionally, the clavicle acts mainly as a point of muscle attachment. Some of the literature suggests that with good repair of the muscle, the only functional consequences of surgical removal of the clavicle are limitations in heavy overhead activity39,40 and that its function as a strut41 is therefore less important. This concept seems to be supported by the relatively good function of persons with congenital absence of the clavicle.42 However, others have found that sudden loss of the clavicle in adulthood has a devastating effect on shoulder function.
Important relationships to the clavicle are the subclavian vein and artery and the brachial plexus posteriorly. In fact, the medial anterior curve is often described as an accommodation for these structures and does not form in Sprengel’s deformity, a condition in which the scapula does not descend. Therefore, the attached clavicle does not need to accommodate.43–45 The curve is a landmark for finding the subclavian vein.46 This relationship is more a factor in surgery than in trauma because the bone acts as an obstruction to surgeons in reaching the nerve or vessel tissue that they wish to treat. In trauma, clavicular injury usually does not affect these structures despite their close relationship, and nonunion is rare.47 Most cases of neurovascular trauma fall into two groups: injury to the carotid artery from the displaced medial clavicle and compression of structures over the first rib.48
Acromioclavicular Joint
The acromioclavicular joint is the only articulation between the clavicle and the scapula, although a few persons, as many as 1%, have a coracoclavicular bar or joint.49,50 Lewis50 reported in his work that about 30% of cadavers had articular cartilage on their opposing coracoid and clavicular surfaces, without a bony process on the clavicle directed toward the coracoid.
The capsule of the acromioclavicular joint contains a diarthrodial joint incompletely divided by a disk, which, unlike that of the sternoclavicular joint, usually has a large perforation in its center.29,30 The capsule tends to be thicker on its superior, anterior, and posterior surfaces than on the inferior surface. The upward and downward movement allows rotation of about 20 degrees between the acromion and the clavicle, which occurs in the first 20 and last 40 degrees of elevation.20 It is estimated that many persons have even less range of motion because in some cases, fusion of the acromioclavicular joint does not decrease shoulder motion.22 DePalma found degenerative changes of both the disk and articular cartilage to be the rule rather than the exception in specimens in the fourth decade or older.29
The blood supply to the acromioclavicular joint is derived mainly from the acromial artery, a branch of the deltoid artery of the thoracoacromial axis. There are rich anastomoses between this artery, the suprascapular artery, and the posterior humeral circumflex artery. The acromial artery comes off the thoracoacromial axis anterior to the clavipectoral fascia and perforates back through the clavipectoral fascia to supply the joint. It also sends branches anteriorly up onto the acromion. Innervation of the joint is supplied by the lateral pectoral, axillary, and suprascapular nerves.27
The ligaments about the acromioclavicular articulation and the trapezoid and conoid ligaments have been studied extensively (Fig. 2-12). Traditionally and more recently, it has been reported that anteroposterior stability of the acromioclavicular joint was controlled by the acromioclavicular ligaments and that vertical stability was controlled by the coracoclavicular ligaments.22,51 A serial cutting experiment involving 12 force-displacement measurements was performed with more sophisticated equipment.52 Three anatomic axes of the acromioclavicular joint were used, and translation and rotation on each axis in both directions were measured. The results of the experiment confirmed previously held views, particularly when displacements were large.
Interpretation of the stability attributed to the acromioclavicular ligaments needs to reflect the additional role that they play in maintaining integrity of the acromioclavicular joint. Although we would expect the linear arrangement of the collagen of the acromioclavicular ligaments to resist distraction, it makes little sense that the acromioclavicular ligaments would resist compression with these fibers, yet 12% to 16% of compression stability in the study was attributed to the acromioclavicular ligament. Maintenance of the integrity of the acromioclavicular joint, particularly the position of the interarticular disk, might be the explanation. We would not expect the acromioclavicular ligament to resist superior translation of the clavicle were it not for the presence of an intact joint below it creating a fulcrum against which these ligaments can produce a tension band effect.52
It is seldom that these ligaments are called on to resist trauma, and their usual function is to control joint motion. As noted earlier, this joint has relatively little motion, and muscles controlling scapulothoracic motion insert on the scapula. To a large extent, the ligaments function to guide motion of the clavicle.53 For example, the conoid ligament produces much of the superior rotation of the clavicle as the shoulder is elevated in flexion.12
The distal end of the clavicle does not have a physeal plate. Todd and D’Errico,54 using microscopic dissection, found a small fleck of bone in some persons that appeared to be an epiphysis, but it united within 1 year. We have not seen this structure at surgery or by roentgenogram. Probably the articular cartilage functions in longitudinal growth as it does in a physis.
Scapula
The scapula is a thin sheet of bone that functions mainly as a site of muscle attachment (Fig. 2-13). It is thicker at its superior and inferior angles and at its lateral border, where some of the more powerful muscles are attached (Figs. 2-14 and 2-15). It is also thick at sites of formation of its processes: the coracoid, spine, acromion, and glenoid. Because of the protection of overlying soft tissue, fractures usually occur in the processes via indirect trauma. The posterior surface of the scapula and the presence of the spine create the supraspinatus and the infraspinatus fossae. The three processes, the spine, the coracoid, and the glenoid, create two notches in the scapula. The suprascapular notch is at the base of the coracoid, and the spinoglenoid, or greater scapular notch, is at the base of the spine. The coracoacromial and transverse scapular ligaments are two of several ligaments that attach to two parts of the same bone. Sometimes an inferior transverse scapular ligament is found in the spinoglenoid notch. This transverse ligament and ganglia of the labrum might all be factors in suprascapular nerve deficits. Seldom studied is the coracoglenoid ligament, which originates on the coracoid between the coracoacromial and coracohumeral ligaments and inserts on the glenoid near the origin of the long head of the biceps.55 The major ligaments that originate from the scapula are the coracoclavicular, coracoacromial, acromioclavicular, glenohumeral, and coracohumeral.
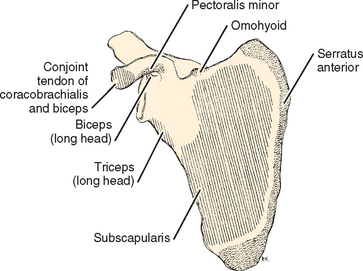
The coracoid process comes off the scapula at the upper base of the neck of the glenoid and passes anteriorly before hooking to a more lateral position. It functions as the origin of the short head of the biceps and the coracobrachialis tendons. It also serves as the insertion of the pectoralis minor muscle and the coracoacromial, coracohumeral, and coracoclavicular ligaments. Several anomalies of the coracoid have been described. As much as 1% of the population has an abnormal connection between the coracoid and the clavicle: a bony bar or articulation.49 Some surgeons have seen impingement in the interval between the head of the humerus and the deep surface of the coracoid.56–58 The coracohumeral interval is smallest in internal rotation and forward flexion.
The spine of the scapula functions as part of the insertion of the trapezius on the scapula and as the origin of the posterior deltoid. It also suspends the acromion in the lateral and anterior directions and thus serves as a prominent lever arm for function of the deltoid. The dimensions of the spine of the scapula are regular, with less than 1.5-cm variation from the mean in any dimension. Recently, reconstruction of the mandible has been devised by using the spine of the scapula. Sacrifice of the entire spine, including the acromion, has a predictably devastating effect on shoulder function.59,60
Because of the amount of pathology involving the acromion and the head of the humerus, the acromion is the most-studied process of the scapula.61 Tendinitis and bursitis have been related to impingement of the head of the humerus and the coracoacromial arch in an area called the supraspinatus outlet.62 When viewed from the front, a 9- to 10-mm gap (6.6-13.8 mm in men, 7.1-11.9 mm in women) can be seen between the acromion and the humerus.63 Recent advances in x-ray positioning allow better visualization of the outlet from the side or sagittal plane of the scapula.64
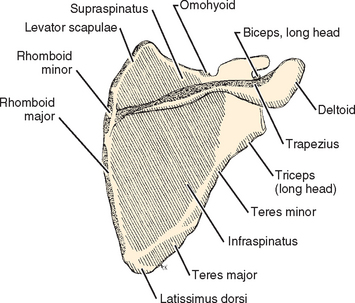
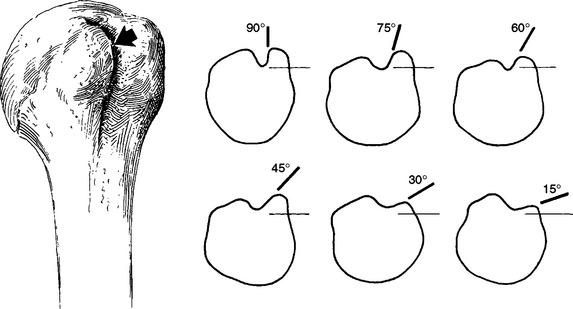
Several methods of describing the capaciousness of this space or its tendency for mechanical discontinuity have been devised. Aoki and colleagues65 used the slope of the ends of the acromion relative to a line connecting the posterior acromion with the tip of the coracoid of the scapula to determine the propensity for impingement problems. Bigliani and associates66 separated acromia into three types (or classes) based on their shape and correlated the occurrence of rotator cuff pathology in cadavers with the shape of the acromion on supraspinatus outlet radiographs (Fig. 2-16). Their classification is generally easy to use, but the transition between types is smooth, so some inter-interpreter variability will occur in those close to the transitions. Type I acromia are those with a flat undersurface and the lowest risk for impingement syndrome and its sequelae. Type II has a curved undersurface, and type III has a hooked undersurface. As one would expect, a type III acromion with its sudden discontinuity in shape had the highest correlation with subacromial pathology.
A more recent report by Banas and associates comments on the position of the acromion in the coronal plane (the lateral downward tilt). In their series of 100 MRI procedures, increasing downward tilt was associated with a greater prevalence of cuff disease.67 The remainder of the roof of the supraspinatus outlet consists of the coracoacromial ligament, which connects two parts of the same bone. It is usually broader at its base on the coracoid, tapers as it approaches the acromion, and has a narrower but still broad insertion on the undersurface of the acromion; it covers a large portion of the anterior undersurface of the acromion and invests the tip and lateral undersurface of the acromion (Fig. 2-17). The ligament might not be wider at its base and often has one or more diaphanous areas at the base.68 Because of the high incidence of impingement in elevation and internal rotation, acromia from persons older than the fifth decade often have secondary changes such as spurs or excrescences.
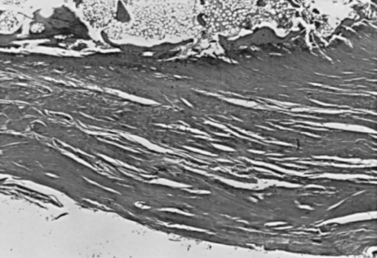
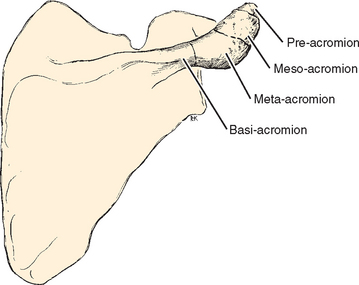
In addition to static deformation of the acromion, one would expect an unfused acromion epiphysis to lead to deformability of the acromion on an active basis and decrease the space of the supraspinatus outlet.69 Neer, however, found no increased incidence of unfused epiphyses in his series of acromioplasties.70 Liberson71 classified the different types of unfused acromia as pre-acromion, meso-acromion, meta-acromion, and basi-acromion centers (Fig. 2-18). In his series, an unfused center was noted on 1.4% of roentgenograms and bilaterally in 62% of cases. The meso-acromion-meta-acromion defect was found most often (Fig. 2-19).
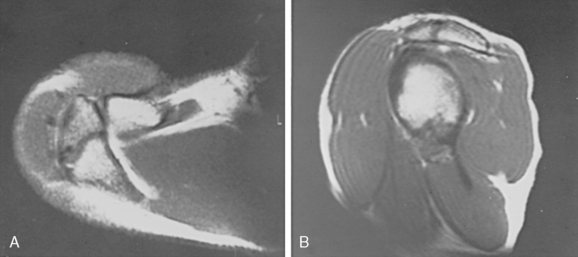
The glenoid articular surface is within 10 degrees of being perpendicular to the blade of the scapula, with the mean being 6 degrees of retroversion.72 The more caudad portions face more anteriorly than the cephalad portions.73 This perpendicular relationship, combined with the complementary orientation of the scapula and relationships determined by the ligaments of the scapulohumeral orientation, makes the plane of the scapula the most suitable coronal plane for physical and radiologic examination of the shoulder. The plane of the glenoid defines the sagittal planes, whereas the transverse plane remains the same.74
The blood supply to the scapula is derived from vessels in muscles that have fleshy origin from the scapula (see the section “Muscles”). Vessels cross these indirect insertions and communicate with bony vessels. The circulation of the scapula is metaphyseal; the periosteal vessels are larger than usual, and they communicate freely with the medullary vessels rather than being limited to the outer third of the cortex. Such anatomy might explain why subperiosteal dissection is bloodier here than over a diaphyseal bone.75 The nutrient artery of the scapula enters into the lateral suprascapular fossa60 or the infrascapular fossa.76 The subscapular, suprascapular, circumflex scapular, and acromial arteries are contributing vessels.
Humerus
The articular surface of the humerus at the shoulder is spheroid, with a radius of curvature of about 2.25 cm.12 As one moves down the humerus in the axis of the spheroid, one encounters a ring of bony attachments for the ligaments and muscles that control joint stability. The ring of attachments is constructed of the two tuberosities, the intertubercular groove, and the medial surface of the neck of the humerus. Ligaments and muscles that maintain glenohumeral stability do so by contouring the humeral head so that tension in them produces a restraining force toward the center of the joint (Fig. 2-20). In this position, the spheroid is always more prominent than the ligamentous or muscle attachments. For example, when the shoulder is in neutral abduction and the supraspinatus comes into play, the greater tuberosity, which is the attachment of this tendon, is on average 8 mm less prominent than the articular surface, and thus the tendon contours the humeral head.77 In the abduction and external rotation position, contouring of the supraspinatus is lost. The anterior inferior glenohumeral ligament now maintains joint stability, and its attachments are less prominent than the articulating surface.
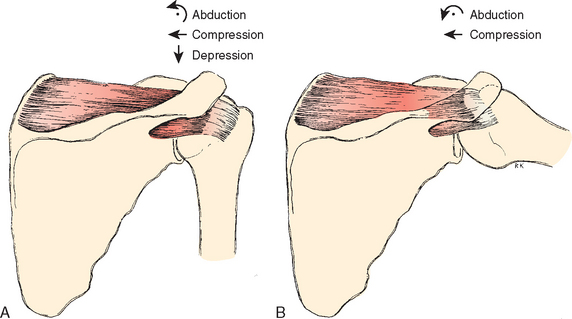
With the arm in the anatomic position (i.e., with the epicondyles of the humerus in the coronal plane), the head of the humerus is retroverted in relation to the transepicondylar axis. In addition, the average retrotorsion is less at birth than at maturity.78 How much retroversion has been a topic of debate. Boileau and Walch79 used three-dimensional computerized modeling of cadaveric specimens to analyze the geometry of the proximal humerus. They found a wide variation of retroversion ranging from −6.7 degrees to 47.5 degrees. These findings have helped revolutionize shoulder arthro-plasty and brought about the development of the third-generation shoulder prosthesis, or the concept of the anatomic shoulder replacement. This concept centers around the great range in retroversion among different people. The surgical goal is restoring individual patient anatomy. Setting prosthetic replacements at an arbitrary 30 to 40 degrees of retroversion might not be optimal and does not account for individual anatomic variability.79 The intertubercular groove lies approximately 1 cm lateral to the midline of the humerus.72,80 The axis of the humeral head crosses the greater tuberosity about 9 mm posterior to the bicipital groove (Fig. 2-21).81 The lesser tubercle (or tuberosity) lies directly anterior, and the greater tuberosity lines up on the lateral side. In the coronal plane the head-to-shaft angle is about 135 degrees.77 This angle is less for smaller heads and greater for larger ones. The head size (radius of curvature) correlates most strongly with the patient’s height.77
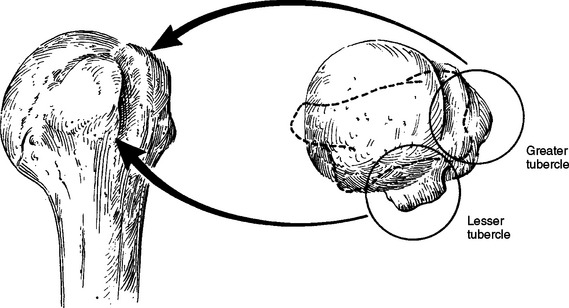
The space between the articular cartilage and the ligamentous and tendon attachments is referred to as the anatomic neck of the humerus (Fig. 2-22). It varies in breadth from about 1 cm on the medial, anterior, and posterior sides of the humerus to essentially undetectable over the superior surface, where no bone is exposed between the edge of articular cartilage and the insertion of the rotator cuff. The lesser tubercle is the insertion for the subscapularis tendon, and the greater tubercle bears the insertion of the supraspinatus, infraspinatus, and teres minor in a superior-to-inferior order. Because of its distance from the center of rotation, the greater tubercle lengthens the lever arm of the supraspinatus as elevation increases above 30 degrees. It also acts as a pulley by increasing the lever arm of the deltoid below 60 degrees.82 The prominence of the greater tubercle can even allow the deltoid to act as a head depressor when the arm is at the side.83 Below the level of the tubercles, the humerus narrows in a region that is referred to as the surgical neck of the humerus because of the common occurrence of fractures at this level.
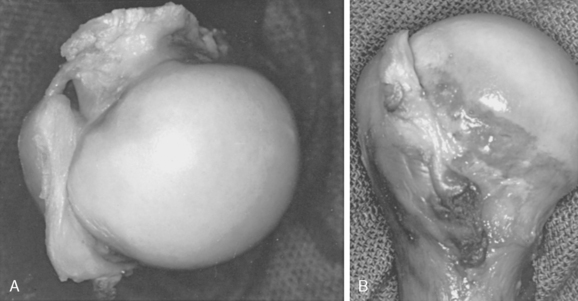
The greater and lesser tubercles make up the boundaries of the intertubercular groove through which the long head of the biceps passes from its origin on the superior lip of the glenoid. The intertubercular groove has a peripheral roof referred to as the intertubercular ligament or transverse humeral ligament, which has varying degrees of strength.58,84 Research has shown that the coracohumeral ligament is the primary restraint to tendon dislocation.85–87 The coracohumeral ligament arises from the coracoid as a V-shaped band, the opening of which is directed posteriorly toward the joint. In most cases, it histologically represents only a V-shaped fold of capsule and has no distinct ligamentous fibers.88 Tightening of this area does affect shoulder function. The ring of tissue making up the pulley constraining the biceps tendon consists of the superficial glenohumeral ligament (floor) and the coracohumeral ligament (roof).
Because the biceps tendon is a common site of shoulder pathology, attempts have been made to correlate the anatomy of its intertubercular groove with a predilection for pathology (Fig. 2-23).27 It was thought that biceps tendinitis resulted from dislocation of the tendon secondary to a shallow groove or a supratubercular ridge84 and an incompetent transverse humeral ligament. Meyer84 attributed the greater number of dislocations of the biceps tendon on the left to activities in which the left arm is in external rotation, a position that should have been protective. Current opinion is that dislocation of the tendon is a relatively rare etiology of bicipital tendinitis, that most cases of bicipital tendinitis can be attributed to impingement,63 and that dislocation of the tendon is not seen except in the presence of rotator cuff or pulley damage.
Walch and colleagues89 analyzed long head of biceps dislocations. They found that in 70% of cases, dislocation of the long head of the biceps was associated with massive rotator cuff tears. In particular, in only 2 of 46 cases was the subscapularis intact. It is possible that the theory of variable depth of the intertubercular groove also applies to the impingement syndrome as an etiology. A shallow intertubercular groove makes the tendon of the long head of the biceps and its overlying ligaments more prominent and therefore more vulnerable to impingement damage.70
The intertubercular groove has a more shallow structure as it continues distally, but its boundaries, referred to as the lips of the intertubercular groove, continue to function as sites for muscle insertion. Below the subscapularis muscles, the medial lip of the intertubercular groove is the site of insertion for the latissimus dorsi and teres major, with the latissimus dorsi insertion being anterior, often on the floor of the groove. The pectoralis major has its site of insertion at the same level but on the lateral lip of the bicipital groove. At its upper end, the intertubercular groove also functions as the site of entry of the major blood supply of the humeral head, the ascending branch of the anterior humeral circumflex artery, which enters the bone at the top of the intertubercular groove or one of the adjacent tubercles.90,91
The humerus, as part of the peripheral skeleton, is rarely a barrier to dissection. The essential relationships to be maintained in surgical reconstruction are the retrograde direction of the articular surface and this surface’s prominence relative to the muscle and ligamentous attachments. Longitudinal alignment needs to be maintained, as well as the distance from the head to the deltoid insertion. In fractures above the insertion of the deltoid that heal in humerus varus, or in cases of birth injury that cause humerus varus, the head-depressing effect of the supraspinatus is ineffective in the neutral position when the shear forces produced by the deltoid are maximal.92 Interestingly, patients with congenital humerus varus rarely complain of pain but have limitation of motion.93 The important relationships to the humerus in the region of the joint are the brachial plexus structures, particularly the axillary and radial nerves and the accompanying vessels.
MUSCLES
The orthopaedic surgeon views muscles in several ways. First, they produce force. Second, they are objects of dissection in terms of repairing, transferring, or bypassing them on the way to a deeper and closely related structure. Finally, muscles are energy-consuming and controllable organs for which blood and nerve supply must be maintained.
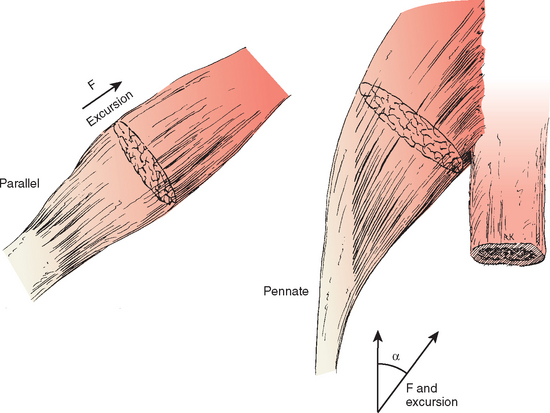
The internal arrangement of muscle fibers can affect strength (Fig. 2-24). If all the fibers of a muscle are arranged parallel to its long axis, the muscle is called parallel and has maximal speed and excursion for size. Other muscles sacrifice this excursion and speed for strength by stacking a large number of fibers in an arrangement oblique to the long axis of the muscle and attaching to a tendon running the length of the muscle. This type of arrangement is referred to as pennate or multipennate. Strength again is a product of the number of fibers; however, because of the oblique arrangement of the fibers, the strength in line with the tendon is obtained by multiplying this strength by the cosine of the angle of incidence with the desired axis of pull. Its excursion is also a product of this cosine times the length of these shorter fibers. Some muscles, such as the subscapularis in its upper portion, have multipennate portions in which excursion is so short and the collagen framework so dense that the muscle acts as a passive restraint to external rotation of the glenohumeral joint (e.g., at 0 and 45 degrees of abduction).94–96
The complex arrangement of the shoulder into several articulations that contribute to an overall increase in mobility is also important in terms of the function of muscle. First, the multiple joint arrangement demands less excursion of the muscles that cross each articulation, thereby allowing each joint’s muscles to operate within the more effective portions of their length-tension curves. Second, joint stability requires that joint reaction forces cross the joint through the bony portions. Muscles of the shoulder serve not only to bring joint reaction forces to the glenohumeral joint in the area of the glenoid but also to move the glenoid to meet the joint reaction forces.97 This need for multiple muscles to function in each motion complicates the diagnosis and planning of tendon transfer.98
Actions of muscles have been measured in terms of standard movements such as abduction in the plane of the scapula, forward flexion of the shoulder, and internal and external rotation of the shoulder in neutral abduction. Studies have also been done on rehabilitation-associated activities such as scapular depression, an activity performed by patients using a wheelchair or walker. More complex motions of daily life and the actions of athletes are currently under research.99 Because the upper extremity can be positioned many ways, an infinite number of studies can be performed.
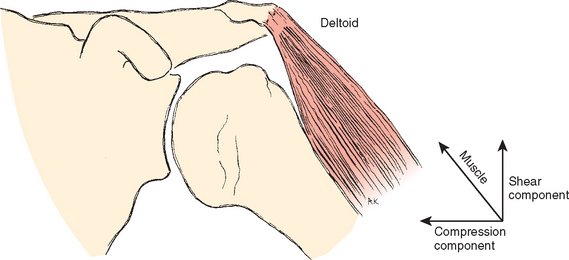
A muscle performs its activity entirely by shortening its length. The action that results and the level of each muscle’s activity depend on two conditions: the position at the beginning of an activity and the relationship of the muscle to the joint. The muscle vectors of the shoulder can be divided into two components with respect to the joint surface (Fig. 2-25). One component produces shear on the joint surface, and the other produces compression and increases the stability of the joint. Muscles of the shoulder can make an additional contribution to stability by circumscribing the protruding portions of the humeral spheroid, a continually directed shear force creating compression to increase stability.12,100 Stability depends on the joint reaction force vector crossing the joint through cartilage and bone, the tissues designed for compression stress. A useful oversimplification of the contributions of shoulder muscles to stability is that the glenohumeral muscles strive to direct the joint reaction vector toward the glenoid and the muscles controlling the scapula move the glenoid toward the force.
Externally applied resistance, including the force of gravity on the action that is being performed, is an additional determinant of which muscles will be active in a particular motion. At high levels of performance, the level of training becomes important in determining how a muscle is used. People who are highly skilled may be more adept than the less trained at positioning their skeletons so that they use less muscle force in retaining joint stability. Such positioning allows more muscle force to be used in moving the limb.101
Two examples illustrate the variability of muscle action in performing the same activity. The first is elevation of the scapula (Fig. 2-26). When the subject is in the anatomic position and is performing an elevation in the scapular plane or in forward flexion, the main elevators of the scapula are the upper fibers of the trapezius, the serratus anterior, and the levator scapulae. In studies of overhead activity102 it was found that the orientation of the throwing arm relative to the spine in different activities such as tennis, baseball, and so forth was similar, but that the arm in relation to the ground was more vertical for heavier objects. The relationship of the humerus to the scapula and the scapula to the main axial skeleton, however, was little changed. Such a finding ought to be anticipated because these muscles are already operating within the optimal portion of Blix’s curve for their activity. Further arm elevation is obtained by using the contralateral trunk muscles, with the upper part of the trunk bending away from the throwing arm and creating a more vertical alignment of the throwing arm relative to the ground. Rotation of the scapula also begins earlier when elevating heavier objects.103 By producing a more level glenoid platform, this maneuver allows the glenoid rather than the muscles to bear more of the weight of the thrown object. In a sense, one might consider the contralateral trunk muscles to be elevators of the scapula.
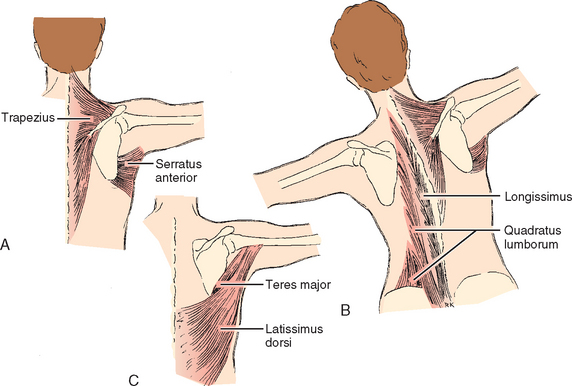
In another athletic maneuver, the iron cross performed by some gymnasts, the arm is again in 90 degrees of abduction, and the entire weight is placed on the hands. The upward force on the hands is counterbalanced by the adduction force generated by the latissimus dorsi and the lower portion of the pectoralis major. Because of the shorter lever arm of these muscles, balancing forces are much greater than the upward forces on the hands (see Chapter 6). Therefore, the joint reaction force on the humeral side tends to run in a cephalocaudad direction. The other muscle active in this enforced adduction is the teres major, which brings the lower portion of the scapula toward the humerus. In this maneuver, the hands are in a fixed position and the teres major acts as an upward rotator of the scapula. Because the teres major is active only against strong resistance,104 it could be postulated that elevation of the scapula may be the major action of the muscle.
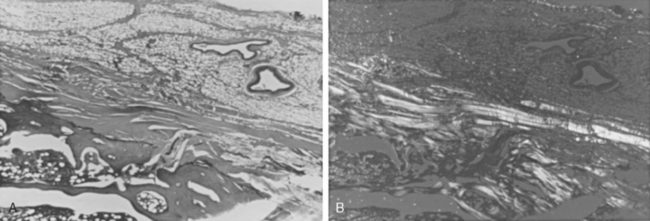
Tendons are attached to bone by interlocking of the collagen of the tendon with the surface of the bone (direct insertion) or by continuation of the majority of collagen fibers into the periosteum (indirect insertion) (Fig. 2-27).105 Direct insertions have a transition zone from the strong but pliable collagen of the tendon to the hard and unyielding calcified collagen of the bone. The transition begins on the tendon side with nonmineralized fibrocartilage, then mineralized fibrocartilage, and finally bone. Thus, there is no sudden transition in material properties at bone attachments, and collagen is present in amounts necessary to bear the stresses generated.
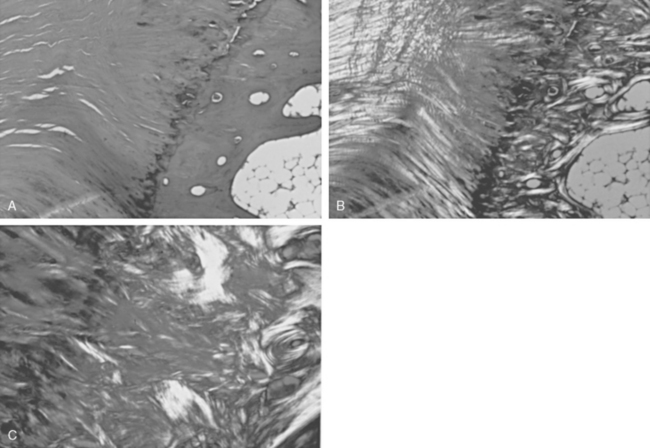
When all the force generated by a muscle is borne by a single narrow tendon, this attachment tends to become a collagen-rich direct insertion and provides the surgeon with a firm structure to hold sutures. When the area of attachment to bone is broad, the same layered arrangement can exist, but most of the collagen goes into periosteum in what is called indirect attachment. As the muscle force is spread over a broad area, these attachments tend to become collagen poor, or fleshy, with little collagen to hold sutures. When these areas of muscle are mobilized, a portion of contiguous periosteum is left attached to the muscle to increase the amount of collagen available to hold sutures during reattachment. Direct insertions are not traversed by blood vessels, but indirect insertions are (Fig. 2-28).105

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


