CHAPTER 1 Developmental Anatomy of the Shoulder and Anatomy of the Glenohumeral Joint
As humans evolved to assume an orthograde posture, the scapulohumeral complex underwent changes to facilitate prehension and comply with the demands of a non-weight-bearing joint. Over time, the inherent osseous articular congruity of the upper limbs was sacrificed for soft tissue stability to achieve a greater degree of mobility at the glenohumeral joint.
COMPARATIVE ANATOMY
General Development
The forelimb in humans is a paired appendage whose evolutionary roots can be traced to the longitudinal lateral folds of epidermis in the fish species Rhipidistian crossopterygian.1 These folds extend caudad from the region just behind the gills to the anus (Fig. 1-1). The pectoral and pelvic fins developed from the proximal and distal portions respectively and were the predecessors of the human upper and lower limbs (Fig. 1-2).2
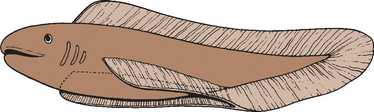
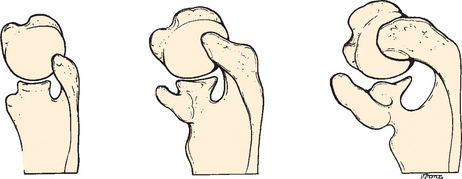
Cartilage rays called radials (Fig. 1-3) arose between muscle buds to form a support structure, and the proximal portions of these radials coalesced to form basal cartilage, or basilia. The radials began to fuse at their base and eventually formed a concrescent central axis, or pectoral girdle (Fig. 1-4). These paired basilia eventually migrated ventrally toward the midline anteriorly to form a ventral bar, which corresponds to the paired clavicles in some mammals, as well as the cleitrum, a membranous bone that attached the pectoral girdle to the skull. The basilia also projected dorsally over the thorax to form the precursor of the scapula. Articulations within the basilia eventually developed at the junction of the ventral and dorsal segments (glenoid fossa) with the remainder of the pectoral fin, which corresponds to the glenohumeral joint in humans (Fig. 1-5).

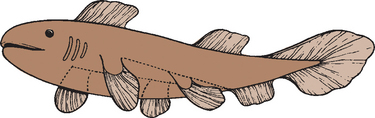
As these prehistoric fish evolved into amphibians, their osseous morphology also changed to adapt to waterless gravity. The head was eventually freed from its attachments to the pectoral girdle, and in the reptile, the pectoral girdle migrated a considerable distance caudally.3 The pelycosaurus of the late Paleozoic Era (235-255 million years ago) is among the oldest reptiles believed to have been solely land dwellers.4 These early tetrapods ambulated with the proximal part of their forelimbs held in the horizontal plane and distal part flexed at a 90-degree angle in the sagittal plane. Locomotion was attained by rotation of the humerus in its longitudinal axis. The cleitrum disappeared entirely in this reptilian stage.
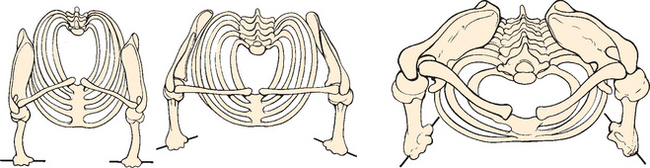
Whereas structural stability was primarily achieved via osseous congruity in these early reptiles, the shoulder evolved to dispense more flexibility and mobility in subsequent species. The basic mammalian pattern developed with articulations arising between a well-developed clavicle and sternum medially and a flat, fairly wide scapula laterally. The coracoid enlarged during this period, and the scapular spine developed in response to new functional demands (Fig. 1-6). Four main variations on this scheme are seen.5 Mammals adapted for running have lost their clavicle to further mobilize the scapula, and the scapula is relatively narrowed. Mammals adapted for swimming also have lost the clavicle, although the scapula is wider and permits more varied function. Shoulder girdles modified for flying have a large, long, well-developed clavicle with a small, narrow, curved scapula. Finally, shoulders modified for brachiating (including those of humans) developed a strong clavicle, a large coracoid, and a widened, strong scapula.
Other adaptations in the erect posture were relative flattening of the thorax in the anteroposterior dimension, with the scapula left approximately 45 degrees to the midline (Fig. 1-7), and evolution of the pentadactyl limb with a strong, mobile thumb and four ulnar digits. This pentadactyl limb is very similar to the human arm as we know it.
In approaching the more human form, we now discuss evolution of the different regions of the shoulder and pectoral girdle separately.
Development of Individual Regions
The Scapula
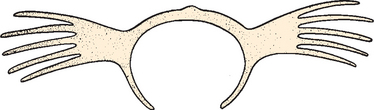
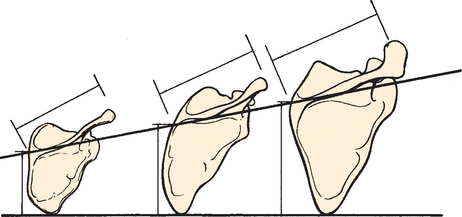
The scapula in humans is suspended by muscles alone and clearly reflects the adaptive development of the shoulder. It has shifted caudally from the cervical position in lower animals, and as a result, the shoulder is freed from the head and neck and can serve as a base or platform to facilitate arm movement. The most striking modification in the development of the bone of the scapula itself is in the relationship between the length (measured along the base of the spine) and the breadth (measured from the superior to the inferior angle) of the scapula, or the scapular index (Fig. 1-8).6 This index is extremely high in the pronograde animal with a long, narrow scapula. In primates and humans, the scapula broadens, and the most pronounced changes are confined to the infraspinatus fossa. This modification has been referred to as an increase in the infraspinatus index.
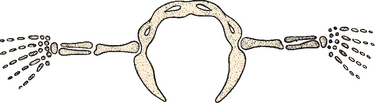
Broadening of the infraspinatus fossa results in a change in the vector of muscle pull from the axillary border of the scapula to the glenoid fossa and consequently alters the action of the attached musculature. This adaptation allows the infraspinatus and teres minor to be more effective in their roles as depressors and external rotators of the humeral head. The supraspinatus fossa and muscle have changed little in size or shape over time; the acromion, which is an extension of the spine of the scapula (see Fig. 1-6), has enlarged over time. In pronograde animals, the acromion process is insignificant; in humans, however, it is a massive structure overlying the humeral head. This change reflects the increasing role of the deltoid muscle in shoulder function. By broadening its attachment on the acromion and shifting its insertion distally on the humerus, it increases its mechanical advantage in shoulder motion.
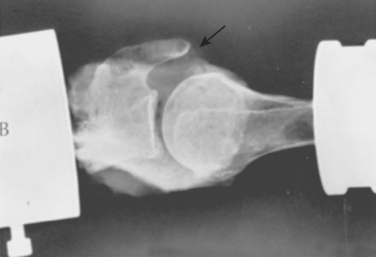
The coracoid process has also undergone an increase in size over time (see Fig. 1-6).6 We have performed biomechanical studies in which it was shown that with the shoulder in 90 degrees of abduction, the coracoid extension over the glenohumeral joint can mechanically limit anterior translation of the humerus relative to the glenoid. In one shoulder that we tested after sectioning of the capsule, the shoulder would not dislocate anteriorly in full abduction until after the coracoid process was removed (Fig. 1-9).7
Humerus
Like the scapula, the humerus has undergone several morphologic changes during its evolution. The head of the humerus has moved proximally, underneath the torso, as well as from the horizontal plane to a more vertical resting orientation. The insertion site of the deltoid has migrated distally to improve the lever arm of the deltoid muscle (Fig. 1-10).6,8
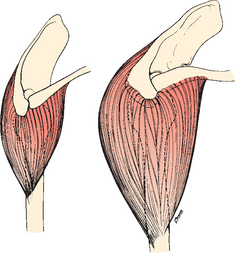
FIGURE 1-10 The deltoid muscle has migrated distally over time to improve the lever arm on the humerus.
In addition, the distal humeral shaft underwent an episode of torsion relative to the proximal end of the humerus, thereby making the humeral head internally rotated relative to the epicondyles.6 As the thoracic cage flattened in the anteroposterior plane, the scapula and glenoid fossa assumed a more dorsal position in the thoracic cage, which led to the glenoid fossa being directed more laterally (see Fig. 1-7). As a consequence, external rotation of the humeral head and internal rotation of the shaft relative to it occurred and led to medial displacement of the intertubercular groove and decreased size of the lesser tuberosity relative to the greater tuberosity. The resultant retroversion of the humeral head has been reported to be 33 degrees in the dominant shoulder and 29 degrees in the nondominant shoulder relative to the epicondyles of the elbow in the coronal plane.9
Scapulohumeral Muscles
The scapulohumeral muscles include the supraspinatus, infraspinatus, teres minor, subscapularis, deltoid, and teres major. The supraspinatus has remained relatively static morphologically but has progressively decreased in relative mass (Fig. 1-11).8 The deltoid, on the other hand, has more than doubled in proportional representation and constitutes approximately 41% of the scapulohumeral muscle mass. This increase in size also increases the overall strength of the deltoid. In lower animals, a portion of the deltoid attaches to the inferior angle of the scapula. In humans, these fibers correspond to the teres minor muscle and explain the identical innervation in these two muscles by the axillary nerve.
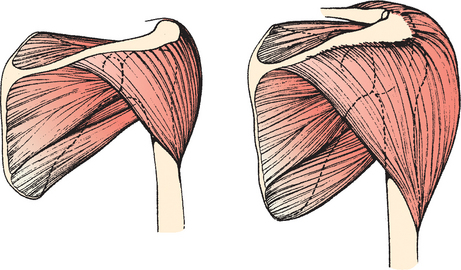
Axioscapular Muscles
The axioscapular muscles include the serratus anterior, rhomboids, levator scapulae, and trapezius. All these muscles (except the trapezius) originated from one complex of muscle fibers arising from the first eight ribs and the transverse processes of the cervical vertebrae and inserting into the vertebral border of the scapula. As differentiation occurred, the fibers concerned with dorsal scapular motion became the rhomboid muscles. The fibers controlling ventral motion developed into the serratus anterior muscle. Finally, the levator scapulae differentiated to control cranial displacement of the scapula. The trapezius has undergone little morphologic change throughout primate development.
This group of muscles acts to anchor the scapula on the thoracic cage while allowing freedom of motion. Most authorities report the ratio between glenohumeral and scapulothoracic motion to be 2:1.6,10 The serratus anterior provides horizontal stability and prevents winging of the scapula.
EMBRYOLOGY
Prenatal Development
The embryonic period is important because all the major external and internal organs develop during this time, and by the end of this period, differentiation is practically complete. All the bones and joints have the form and arrangement characteristic of adults. Exposure to teratogens during this period can cause major congeni tal malformations. During the fetal period, the limbs grow and mature as a result of a continual remodeling and reconstructive process that enables a bone to maintain its characteristic shape. In the skeleton in general, increments of growth in individual bones are in precise relationship to those of the skeleton as a whole. Ligaments show an increase in collagen content, bursae develop, tendinous attachments shift to accommodate growth, and epiphyseal cartilage becomes vascularized.
Few studies have focused on prenatal development of the glenohumeral joint. The contributions by DePalma and Gardner were essential but did not emphasize clinical correlations between the observed fetal anatomy and pathology seen in the postnatal shoulder.11–13 Most studies of the developing shoulder have focused primarily on bone maturation. Analysis of soft tissue structures of the developing shoulder, such as the joint capsule and the labrum, is still incomplete. Studies have not thoroughly evaluated the inferior glenohumeral ligament complex, which has been shown to be an integral component for stability in the adult.14 The seminal studies of the fetal glenohumeral joint were completed before the role of the soft tissue structures in shoulder stability was elucidated. We now have a greater appreciation of the anatomy and biomechanics of the static and dynamic stabilizers of the glenohumeral joint and their role in shoulder stability.
Embryonic Period
The limb buds are initially seen as small elevations on the ventrolateral body wall at the end of the fourth week of gestation.15 The upper limb buds appear during the first few days and maintain a growth advantage over the lower limbs throughout development. Because development of the head and neck occurs in advance of the rest of the embryo, the upper limb buds appear disproportionately low on the embryo’s trunk (Fig. 1-12). During the early stages of limb development, the upper and lower extremities develop in similar fashion, with the upper limb bud developing opposite the lower six cervical and the first and second thoracic segments.
At 4 weeks, the upper limb is a sac of ectoderm filled with mesoderm and is approximately 3 mm long. Each limb bud is delineated dorsally by a sulcus and ventrally by a pit. The pit for the upper limb bud is called the fossa axillaris. The mesoderm in the upper limb bud develops from somatic mesoderm and consists of a mass of mesenchyme, which is loosely organized embryonic connective tissue. Mesenchymal cells can differentiate into many different cells, including fibroblasts, chondroblasts, and osteoblasts (Fig. 1-13). Most bones first appear as condensations of these mesenchymal cells, from which a core called the blastema is formed.15,16 This development is orchestrated by the apical ectodermal ridge (Fig. 1-14), which exerts an inductive influence on the limb mesenchyme, promoting growth and development.
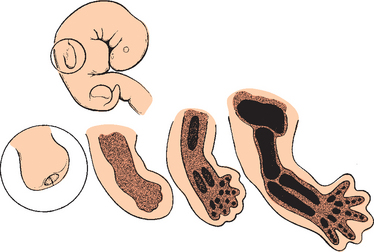
FIGURE 1-14 The apical ectodermal ridge exerts an inductive influence on the development of the upper limb.
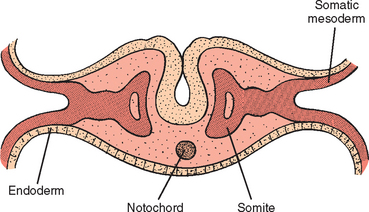
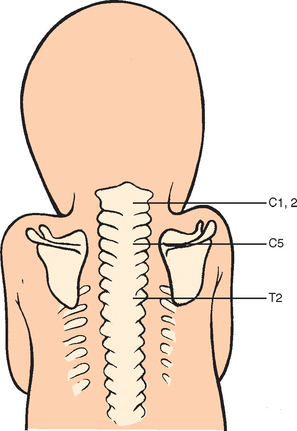
During the fifth week, a number of developments occur simultaneously. The peripheral nerves grow from the brachial plexus into the mesenchyme of the limb buds. Such growth stimulates development of the limb musculature, where in situ somatic limb mesoderm aggregates and differentiates into myoblasts and discrete muscle units. This process is different from development of the axial musculature, which arises from the myotomic regions of somites, or segments of two longitudinal columns of paraxial mesoderm (Fig. 1-15). Also at this time, the central core of the humerus begins to chondrify, although the shoulder joint is not yet formed. There is an area in the blastema called the interzone that does not undergo chondrification and is the precursor of the shoulder joint (Fig. 1-16). The scapula at this time lies at the level of C4 and C5 (Fig. 1-17),17 and the clavicle is beginning to ossify (along with the mandible, the clavicle is the first bone to begin to ossify).
During the sixth week, the mesenchymal tissue in the periphery of the hand plates condenses to form digital rays. The mesodermal cells of the limb bud rearrange themselves to form a deep layer, an intermediate layer, and a superficial layer. This layering is brought on by differential growth rates.18 Such differential growth in the limb also stimulates bending at the elbow because the cells on the ventral side grow faster than those on the dorsal side, which stretches to accommodate the ventral growth. The muscle groups divide into dorsal extensors and ventral flexors, and the individual muscles migrate caudally as the limb bud develops. In the shoulder joint, the interzone assumes a three-layered configuration, with a chondrogenic layer on either side of a loose layer of cells.19 At this time, the glenoid lip is discernible (Fig. 1-18), although cavitation or joint formation has not occurred. Initial bone formation begins in the primary ossification center of the humerus. The scapula at this time undergoes marked enlargement and extends from C4 to approximately T7.
Early in the seventh week, the limbs extend ventrally and the upper and lower limb buds rotate in opposite directions (Fig. 1-19). The upper limbs rotate laterally through 90 degrees on their longitudinal axes, with the elbow facing posteriorly and the extensor muscles facing laterally and posteriorly.15 The lower limbs rotate medially through almost 90 degrees, with the knee and extensor musculature facing anteriorly. The final result is that the radius is in a lateral position in the upper limb and the tibia is in a medial position in the lower limb, although they are homologous bones. The ulna and fibula are also homologous bones, and the thumb and great toe are homologous digits. The shoulder joint is now well formed, and the middle zone of the three-layered interzone becomes less and less dense with increasing cavitation (Fig. 1-20). The scapula has now descended and spans from just below the level of the first rib to the level of the fifth rib.20 The brachial plexus has also migrated caudally and lies over the first rib. The final few degrees of downward displacement of the scapula occur later when the anterior portion of the rib cage drops obliquely downward.
By the eighth week the embryo is about 25 to 31 mm long, and through growth of the upper limb, the hands are stretched with the arms pronated (Fig. 1-21). The musculature of the limb is now also clearly defined. The shoulder joint has the form of the adult glenohumeral joint, and the glenohumeral ligaments can now be visualized as thickenings in the shoulder capsule.15,21
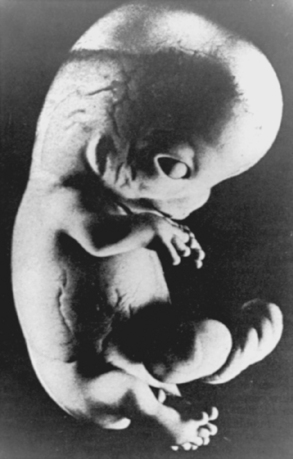
Although certain toxins and other environmental factors can still cause limb deformities (e.g., affecting the vascular supply), it is the embryonic period that is most vulnerable to congenital malformations, with the type of abnormality depending on the time at which the orderly sequence of differentiation was interrupted. One important factor in gross limb abnormalities, such as amelia, involves injury to the apical ectodermal ridge, which has a strong inductive influence on the limb mesoderm. Matsuoka and colleagues have mapped the destinations of embryonic neural crest and mesodermal stem cells in the neck and shoulder region using Cre recombinase-mediated transgenesis.22 A precise code of connectivity that mesenchymal stem cells of both neural crest and mesodermal origin obey as they form muscle scaffolds was proposed. The conclusions suggested that knowledge of these relations could contribute further to identifying the etiology of diseases such as Klippel-Feil syndrome, Sprengel’s deformity, and Arnold-Chiari I/II malformation.22 Clearly, the timing of embryologic development is critical for understanding anomalies and malformations and is an area of further study.
Fetal Period
Fetal development is concerned mainly with expansion in size of the structures differentiated and developed during the embryonic period. By the end of the 12th week, the upper limbs have almost reached their final length. Ossification proceeds rapidly during this period, especially during the 13th to 16th weeks. The first indication of ossification in the cartilaginous model of a long bone is visible near the center of the shaft. Primary centers appear at different times in different bones, but usually between the 7th and 12th weeks. The part of the bone ossified from the primary center is called the di-aphysis. Secondary centers of ossification form the epi- physis. The physeal plate separates these two centers of ossification until the bone grows to its adult length. From the 12th to the 16th week, the epiphyses are invaded by a vascular network, and in the shoulder joint, the epiphysis and part of the metaphysis are intracapsular. The tendons, ligaments, and joint capsule around the shoulder are also penetrated by a rich vascular network during the same time in the fetal period, that is, the third to fourth month of gestation.
A morphologic study of the prenatal developing shoulder joint concluded that the most important changes take place around the 12th week of prenatal life.23 At about this time the glenoid labrum, the biceps tendon, and the glenohumeral ligaments formed a complete ring around the glenoid fossa and led the authors to believe that these structures play a role in stabilizing the joint as well as increasing the concavity of the glenoid fossa. The glenoid labrum consists of dense fibrous tissue and some elastic tissue but no fibrocartilage (as seen in the meniscus of the knee). The acromioclavicular joint develops in a manner different from that of the shoulder joint. Its development begins well into the fetal period (not the embryonic period), and a three-layered interzone is not seen as it is in the glenohumeral joint (Fig. 1-22). Most of the bursae of the shoulder, including the subdeltoid, subcoracoid, and subscapularis bursae, also develop during this time.
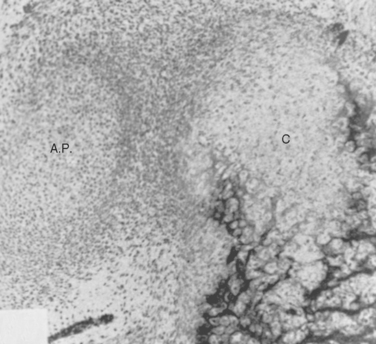
Fealy and colleagues studied 51 fetal glenohumeral joints from 37 specimens to evaluate shoulder morphology on a gross and histologic level and compare it with known postnatal anatomic and clinical findings in fetuses from 9 to 40 weeks of gestation.24 Specimens were studied under a dissecting microscope, histologically, and with the aid of high-resolution radiographs to evaluate the presence of ossification centers. Fetal gross anatomy and morphology were similar to that of normal postnatal shoulders in all specimens. As noted previously, only the clavicle and spine of the scapula were ossified in the fetal shoulder. The humeral head and glenoid gradually and proportionally increased in size with gestational age. Comparative size ratios were consistent except for the fetal coracoid process, which was noted to be prominent in all specimens (Fig. 1-23).

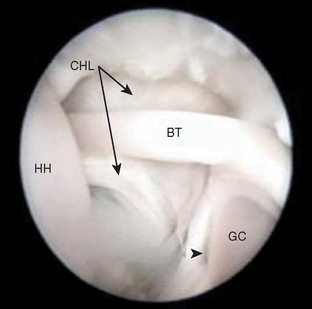
In study by Tena-Arregui and colleagues,25 frozen human fetuses (40 shoulders) were grossly evaluated arthroscopically with similar findings. They concluded that the anatomy observed was easier to discern than what is observed in adult shoulder arthroscopy25 (Fig. 1-24).
Coracoacromial Arch Anatomy
By 13 weeks of gestation, the rotator cuff tendons, coracoacromial ligament (CAL), and coracohumeral ligament are present. The acromion is cartilaginous and consistently has a gentle curve that conforms to the superior aspect of the humeral head, similar to a type II acromion (Fig. 1-25).26–28 These data suggest that variations in acromial morphology are acquired.
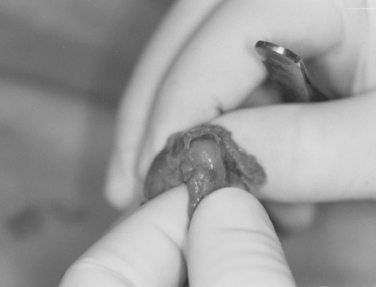
A macroscopic and histologic study performed by Shah and associates analyzed 22 cadaveric shoulders to establish what, if any, developmental changes occur in the differing patterns of acromia.29 In all the curved and hooked acromia (types II and III), a common pattern of degeneration of collagen, fibrocartilage, and bone was observed, consistent with a traction phenomenon. None of these changes were exhibited by the flat acromion (type I). They therefore supported the conclusion that the different shapes of acromion are acquired in response to traction forces applied via the CAL and are not congenital.
The CAL consists of two distinct fiber bundles that lie in the anterolateral and posteromedial planes, as it does in the mature shoulder.30 Histologic studies show that the CAL continues posteriorly along the inferior surface of the anterolateral aspect of the acromion. The CAL has well-organized collagen fiber bundles by 36 weeks of gestation.
In a study by Kopuz and colleagues, 110 shoulders from 60 neonatal cadavers were dissected and analyzed to look for CAL variations.31 Three CAL types were identified: quadrangular, broad band, and V shaped. Histologic analysis showed that V-shaped ligaments had a thin central tissue close to the coracoid. The data suggest that the primordial CAL is broad shaped but assumes a quadrangular shape because of the different growth rates of the coracoid and acromial ends. In addition, broad and V-shaped CALs account for the primordial and quadrangular types, and Y-shaped ligaments account for the adult types of the single- or double-banded anatomic variants, respectively. They concluded that various types of CALs are present during the neonatal period and that the final morphology is determined by developmental factors rather than degenerative changes.
Glenohumeral Capsule and Glenohumeral Ligaments
The anterior glenohumeral capsule was found to be thicker than the posterior capsule. The fetal shoulder capsule inserted onto the humeral neck in the same fashion as in the mature shoulder and was found to be confluent with the rotator cuff tendons at their humeral insertion. Superior and middle glenohumeral ligaments were identifiable as capsular thickenings, whereas the inferior glenohumeral ligament was a distinct structure identifiable by 14 weeks of gestation. Anterior and posterior bands were often noticeable in the ligament, consistent with the known inferior glenohumeral ligament complex (IGHLC) anatomy in the adult shoulder.14 The anterior band of the IGHLC contributed more to formation of the axillary pouch than did the posterior band.
Histologically, the fetal IGHLC consists of several layers of collagen fibers that are highly cellular and have little fibrous tissue during early development. This tissue becomes more fibrous later in gestation. Polarized light microscopy demonstrates that these fibers are only loosely organized but are more organized than adjacent capsular tissues are. Arthroscopic images of the superior glenohumeral ligament have revealed a defined attachment to the humeral head, forming an intersection of the biceps tendon as it enters the bicipital groove and the attachment of the upper edge of the subscapular muscle tendon.25
A rotator interval defect was noted in fetuses by 14 weeks of gestation. This capsular defect was seen consistently in the 1-o’clock position in a right shoulder or the 11-o’clock position in a left shoulder. The interval defect was often covered by a thin layer of capsule that extended from the middle glenohumeral ligament and passed superficially to the defect. Removal of this capsular layer revealed a clear defect between the superior and middle glenohumeral ligaments. Histologic examination of the interval defect in a 19-week-old specimen revealed a thin surrounding capsule with poorly organized collagen fibers. To our knowledge, this is the first suggestion that the capsular defect is not acquired. Specimens with larger rotator interval defects had greater amounts of inferior glenohumeral laxity. Closure of a large rotator interval defect in adults has been shown to be effective treatment of inferior glenohumeral instability.32–34






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


