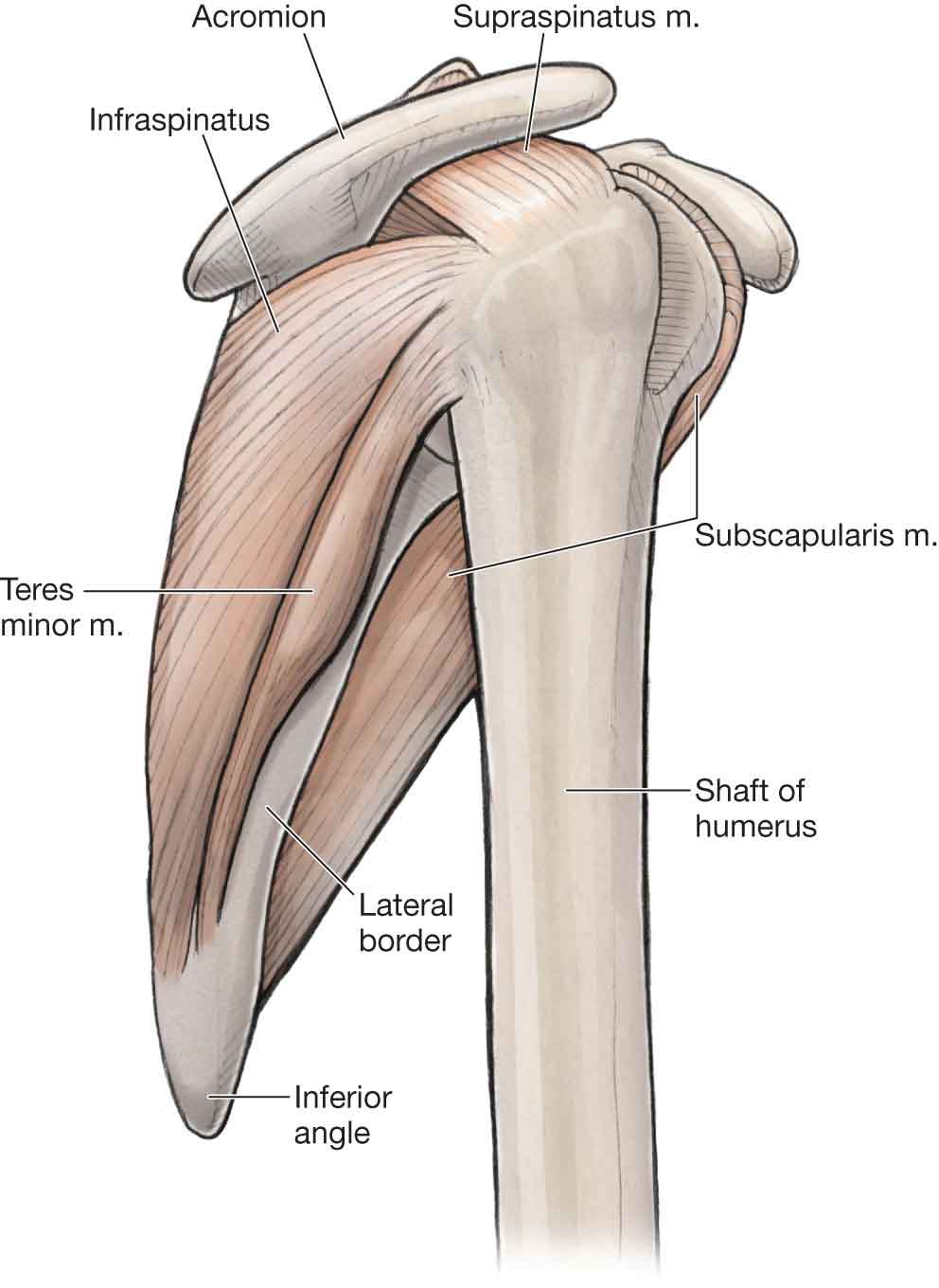
The shoulder is the most rewarding joint in the body because when a limited or painful movement is found, the finding is seldom ambiguous and often implicates the offending structure. —James Cyriax, MD (1904–1985) The primary function of the shoulder complex is to position the hand in space, thereby allowing an individual to interact with his or her environment and to perform fine motor functions. An inability to position the hand results in profound impairment of the entire upper extremity.1 Secondary functions of the shoulder complex include the following: The shoulder is endowed with a unique blend of mobility and stability. Optimal functioning of the shoulder and arm can only take place if a delicate balance between the mobility and stability is maintained. The degree of mobility is contingent on a healthy articular surface, intact muscle–tendon units, and supple capsuloligamentous restraints. The degree of stability is dependent on intact capsuloligamentous structures, proper function of the muscles, and the integrity of the osseous articular structures.1 The shoulder complex functions as an integrated unit, involving a complex relationship between its various components. The components of the shoulder joint complex consist of (Fig. 16-1): FIGURE 16-1 Bony anatomy of the shoulder. (Reproduced, with permission, from Chapter 29. Overview of the Upper Limb. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) For optimal shoulder function, cervicothoracic junction motions, and motions at the connections between the first three ribs and the sternum and spine must be available. The G-H joint is a true synovial joint that connects the upper extremity to the trunk, as part of the upper kinetic chain. The G-H joint is described as a ball and socket joint—the humeral head forms roughly half a ball or sphere (Fig. 16-1), whereas the glenoid fossa forms the socket. The head of the humerus faces medially, posteriorly, and superiorly with the axis of the head forming an angle of 130–150 degrees with the long axis of the humerus (Fig. 16-2).2 In the frontal plane, the head of the humerus is angled posteriorly (retroverted) by 30–40 degrees.3,4 The joint capsule of the G-H joint is lax inferiorly to permit full elevation of the arm. FIGURE 16-2 Superior aspect of shoulder showing angle of scapula. (Reproduced, with permission, from Chapter 30. Shoulder and Axilla. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) The glenoid fossa of the scapula faces laterally, superiorly, and anteriorly at rest and inferiorly and posteriorly when the arm is in the dependent position (Fig. 16-3).5 The glenoid fossa is flat and covers only one-third to one-fourth of the surface area of the humeral head. This arrangement allows for a great deal of mobility but little in the way of articular stability. However, the glenoid fossa is made approximately 50% deeper (doubling the depth of the glenoid fossa across its equatorial line)6 and more concave by a ring of fibrous cartilage7 and dense fibrous collagen8 called a labrum. The labrum forms a part of the articular surface and is attached to the margin of the glenoid cavity and the joint capsule.9 It is also attached to the lateral portion of the biceps anchor superiorly.10 In addition, approximately 50% of the fibers of the long head of the biceps (LHB) brachii originate from the superior labrum (the remainder originates from the superior glenoid tubercle),8 with four different variations identified,11 and continue posteriorly to become a periarticular fiber bundle, making up the bulk of the labrum.10,12 The labrum enhances joint stability by increasing the humeral head contact areas to 75% vertically and 56% transversely.5,9 The humeral–glenoid contact area provides two primary functions:13 it spreads the joint loading over a broad area, and it permits movement of opposing joint surfaces with minimal friction and wear.14 However, because the humeral head is larger than the glenoid (Fig. 16-3), at any point during elevation, only 25–30% of the humeral head is in contact with the glenoid, with the greatest contact occurring during elevation rather than at the extremes. Contact between the humeral head and glenoid fossa is significantly reduced when the humerus is positioned in15–17: FIGURE 16-3 Glenohumeral joint. (Reproduced, with permission, from Chapter 30. Shoulder and Axilla. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) Although the labrum provides some stability for the G-H joint, additional support is provided by both dynamic and static mechanisms. The dynamic mechanisms include the muscles of the rotator cuff (supraspinatus, infraspinatus, teres minor, and subscapularis muscles) (Fig. 16-4) and a number of muscle force couples described later. The static mechanisms, which include reinforcements of the joint capsule, joint cohesion and geometry, and ligamentous support, are also described later. FIGURE 16-4 Rotator cuff muscles. (Reproduced, with permission, from Chapter 30. Shoulder and Axilla. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) The scapula (Fig. 16-1) functions as a stable base from which G-H mobility can occur. The scapula is a flat blade of bone that is oriented to contribute to stability: it lies along the thoracic cage at 30 degrees to the frontal plane (Fig. 16-2), 3 degrees superiorly relative to the transverse plane to augment functional reaching motions above shoulder height,18 and 20 degrees forward in the sagittal plane.19–21 This orientation results in arm elevation occurring in a plane that is 30–45 degrees anterior to the frontal plane. When elevation of the arm occurs in this plane, the motion is referred to as scapular plane abduction or scaption. The scapula’s wide and thin configuration allows for its smooth gliding along the thoracic wall and provides a large surface area for muscle attachments both distally and proximally.24 In all, 16 muscles gain attachment to the scapula (Table 16-1). Six of these muscles, including the trapezius, rhomboids, levator scapulae, and serratus anterior, support and move the scapula, while nine of the other ten (the omohyoid is not included) are concerned with G-H motion.25–27 TABLE 16-1 Muscles of the Scapula Trapezius Subscapularis Levator scapulae Coracobrachialis Long and short head of the biceps Pectoralis minor Rhomboid major Serratus anterior Rhomboid minor Long head of triceps Supraspinatus Teres major Infraspinatus Teres minor Deltoid Omohyoid Posteriorly, the scapula is divided by the elevated scapula spine into two unequally sized muscle compartments. The supraspinous fossa is small and serves as the site of origin for the supraspinatus muscle (see Fig. 16-4). The infraspinous fossa gives attachment for the downward-acting infraspinatus and teres minor muscles (see Fig. 16-4), important muscles for the stabilization of the humeral head (see “Muscles of the Shoulder Complex” section). The spine of the scapula (see Fig. 16-1) provides a continuous line of attachments for the supporting trapezius muscle along its upper border, whereas the deltoid muscle, which suspends the humerus, gains origin from its lower border. The undersurface of the scapula is covered by the subscapularis muscle (see Fig. 16-4), which also assists in stabilizing the humeral head against the glenoid fossa. A prominent feature of the scapula is the large overhanging acromion (see Fig. 16-3), which, along with the coracoacromial ligament and the previously mentioned labrum, functionally enlarges the G-H socket. The position of the acromion also places the deltoid muscle in a dominant position to provide muscular support during elevation of the arm. Bigliani et al.21,28 introduced the following acromion types: The distance, or angle, between the scapula and the clavicle is variable and depends on function. While the shoulder is protracted, the angle is 50 degrees. At rest, the angle is approximately 60 degrees, and with retraction the angle increases to 70 degrees.3 Along the medial border of the scapula arise three muscles: the two rhomboid muscles and the serratus anterior (Fig. 16-5), all of which aid with scapular stability during arm elevation (see later). The coracoid process (see Fig. 16-3) projects forward like a crow’s beak, for which it is named. This forward position provides an efficient lever whereby the small pectoralis muscle can help to stabilize the scapula. In addition, the process serves as a point of origin for the coracobrachialis and the short head of the biceps muscle. FIGURE 16-5 Rhomboid muscles and serratus anterior. (Reproduced, with permission, from Chapter 1. Back. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) The lateral attachment of the voluminous joint capsule of the G-H joint attaches to the anatomical neck of the humerus (Fig. 16-6).6 Medially, the capsule is attached to the periphery of the glenoid and its labrum. The overall strength of the joint capsule bears an inverse relationship with the patient’s age; the older the patient, the weaker the joint capsule. The fibrous portion of the capsule is very lax and has several recesses, depending on the position of the arm. At its inferior aspect, the capsule forms an axillary recess, which is both loose and redundant. The recess permits normal elevation of the arm, although it can also be the site of adhesions when the shoulder is immobilized in adduction. The anterior aspect of the joint capsule is reinforced by three ligaments (Z ligaments), which are described in the next section. The tendons of the rotator cuff (supraspinatus, infraspinatus, teres minor, and subscapularis) reinforce the superior, posterior, and anterior aspects of the capsule, as does the LHB tendon. FIGURE 16-6 Glenohumeral joint and ligaments. (Reproduced, with permission, from Chapter 30. Shoulder and Axilla. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) An inner synovial membrane lines the fibrous capsule and secretes synovial fluid into the joint cavity. The synovium typically lines the joint capsule and extends from the glenoid labrum down to the neck of the humerus. It also forms variously sized bursae, the largest of which, the subacromial or subdeltoid bursa, lies on the superior aspect of the joint (see later). The greater and lesser tuberosities (see Fig. 16-1), which serve as attachment sites for the tendons of the rotator cuff muscles, are located on the lateral aspect of the anatomical neck of the humerus, an imaginary line that separates the humeral head from the rest of the humerus. The lesser tuberosity serves as the attachment for the subscapularis. The greater tuberosity serves as the attachment for the supraspinatus, infraspinatus, and teres minor. The greater and lesser tuberosities are separated by the intertubercular groove (see Fig. 16-1), through which passes the tendon of the LHB on its route to attach to the superior rim of the glenoid fossa. This groove has wide variance in the angle of its walls, but 70% fall within a 60–75-degree range.29 Certain shoulder disorders, including rotator cuff and bicipital tendinopathy, have been associated with anomalies of this groove.30 As the tendon of the LHB (Fig. 16-7) passes over the humeral head from its origin, it makes a right-angle turn to lie in the anterior aspect of the humerus. This abrupt turn may permit abnormal wearing of the tendon at this point. The roof of this groove is formed by the transverse ligament. The transverse humeral ligament (Fig. 16-7), which runs perpendicular over the biceps tendon, was once thought to function as a restraint to the biceps tendon within the intertubercular groove. However, this appears to be the role of the coracohumeral ligament.30 FIGURE 16-7 Soft-tissue structures of the shoulder. (Reproduced, with permission, from Chapter 30. Shoulder and Axilla. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) The region below the greater and lesser tuberosities, where the upper margin of the humerus joins the shaft of the humerus, is referred to as the surgical neck (Fig. 16-1). The axillary nerve and posterior humeral circumflex artery lie in close proximity to the medial aspect of the surgical neck. A number of structures function as static stabilizers during motion of the arm to reciprocally tighten and loosen, thereby limiting translation and rotation of the G-H joint surfaces in a load-sharing fashion.31 These include the G-H ligaments and the posterior capsule. Further static stabilization is provided by the glenoid labrum. The posterior capsule is under tension when the shoulder is in flexion, abduction, IR (in particular, the superior and middle segments), or in any combination of these. At the anterior portion of the outer fibers of the joint capsule, three local reinforcements are present: the superior, middle, and inferior G-H ligaments. Together with the coracohumeral ligament, these ligaments form a Z-pattern on the anterior aspect of the shoulder.32 In the midrange of rotation, the G-H ligaments are relatively lax and stability is maintained primarily by the action of the rotator cuff muscle group compressing the humeral head into the conforming glenoid articulation.31 The G-H ligaments appear to produce a major restraint during shoulder flexion, extension, and rotation33: Other ligaments help provide stability to the G-H joint. These include the following: The coracoacromial arch (Fig. 16-6) is formed by the anteroinferior aspect of the acromion process, and the coracoacromial ligament, which connects the coracoid to the acromion and the inferior surface of the A-C joint.38–40 A number of structures are located in the subacromial or suprahumeral, space (Fig. 16-7) between the coracoacromial arch superiorly and the humeral head inferiorly, and the coracoid process, anteromedially. These include (from inferior to superior) the following: In normal individuals, the subacromial space averages 10–11 mm in height with the arm adducted to the side.41,42 Elevating the arm in the plane of the scapula decreases this space, and the space is at its narrowest between 60 and 120 degrees of scaption.43 During overhead motion in the plane of the scapula, the supraspinatus tendon, the region of the cuff most involved in overuse syndromes of the shoulder, can pass directly underneath the coracoacromial arch. If the arm is elevated while internally rotated, the supraspinatus tendon passes under the coracoacromial ligament, whereas if the arm is externally rotated, the tendon passes under the acromion itself.44 Muscle imbalances or capsular contractures can cause an increase in superior translation of the humeral head, narrowing the subacromial space. For example, if the rotator cuff muscles are weak or injured, increased translation occurs between the humeral head and the glenoid labrum.45 This increase in translation may lead to increased wear on the labrum, increased reliance on the static restraints (e.g., ligaments, capsule), and eccentric overloading of the dynamic (muscle) restraints, which in turn can result in instability and/or a condition termed “subacromial impingement syndrome (SIS).”46,47 Approximately eight bursae are distributed throughout the shoulder complex. The subdeltoid–subacromial bursae (see Fig. 16-7) are usually collectively referred to as the subdeltoid bursa because they are often continuous in nature. The subdeltoid bursa is one of the largest bursae in the body and provides two smooth serosal layers, one of which adheres to the overlying deltoid muscle and the other to the rotator cuff lying beneath. This bursa is also connected to the acromion, greater tuberosity, and coracoacromial ligament. As the humerus elevates, it permits the rotator cuff to slide easily beneath the deltoid muscle. There are also smaller bursae interposed between most of the muscles in contact with the joint capsule: The shoulder complex is embryologically derived from C5 to C8, except the A-C joint, which is derived from C4 (Fig. 16-8). The sympathetic nerve supply to the shoulder originates primarily in the thoracic region from T2 down as far as T8.48 FIGURE 16-8 Neurological and vascular structures of the shoulder. (Reproduced, with permission, from Chapter 30. Shoulder and Axilla. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) One study49 attempted to determine the variability of the course and the pattern of these nerves and found that the peripheral nerves contributing to the anterior shoulder joint included the axillary (C5–6), subscapular (C5–6), and lateral pectoral (C5–6). The same study found that the nerves contributing articular branches to the posterior joint structures are the suprascapular nerve (C5–6) and small branches of the axillary nerve.49 The pathways of these nerves are described in Chapter 3. Other nerves innervate the muscles that act upon the shoulder. These include the long thoracic nerve (C5–7) (Fig. 16-8), which innervates the serratus anterior, the spinal accessory nerve (cranial nerve XI and C3–4), which innervates the sternocleidomastoid (SCM) and trapezius muscles, and the musculocutaneous nerve (C5–7) (Fig. 16-8), which innervates the coracobrachialis, biceps brachii, and brachialis, before dividing into its cutaneous branches. The vascular supply to the shoulder complex is primarily provided by branches off the axillary artery (Fig. 16-9A), which begins at the outer border of the first rib as a continuation of the subclavian artery. The axillary artery is commonly divided into three parts: above, behind, and below the pectoralis minor muscle. The axillary artery meets the more deeply placed brachial plexus in the neck, and here they are encased in the axillary sheath, together with the axillary vein.52 The lateral, posterior, and medial cords of the brachial plexus descend behind the first portion of the artery and then take up their respective locations at the second portion of the artery (behind the pectoralis minor muscle). The branches of these cords also maintain their respective positions. Compression of the axillary artery (or, to a lesser degree, the axillary vein) can result in shoulder dysfunction and most commonly occurs in the posterior fossa around the shoulder and against the humerus with shoulder elevation.52 The G-H joint receives its blood supply from the anterior and posterior circumflex humeral as well as the suprascapular and circumflex scapular vessels (Fig. 16-9B-C).53 The vascular supply to the labrum arises mostly from its peripheral attachment to the capsule and is from a combination of the suprascapular circumflex scapular branch of the subscapular and posterior humeral circumflex arteries (Fig. 16-9C).8 FIGURE 16-9 Vascularization of the shoulder. (Reproduced, with permission, from Chapter 30. Shoulder and Axilla. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) The brachial artery (Fig. 16-9A) provides the dominant arterial supply to each of the two heads of the biceps brachii. The artery travels in the medial intermuscular septum and is bordered by the biceps muscle anteriorly, the brachialis muscle medially, and the medial head of the triceps muscle posteriorly.54 The microvasculature of the rotator cuff has been the subject of much discussion and consists of three main sources: the thoracoacromial, suprahumeral, and subscapular arteries.53 The supraspinatus receives its primary supply from the thoracoacromial arteries. The subscapularis receives its supply from the anterior humeral circumflex (Fig. 16-9A) and the thoracoacromial arteries (Fig. 16-9A). The posterior rotator cuff muscles, the infraspinatus and teres minor, receive their blood supply from the posterior humeral circumflex and suprascapular artery (Fig. 16-9C). The circulation of the rotator cuff is unidirectional with no flow traversing the tide mark at the insertion of the supraspinatus.55 The supraspinatus and biceps tendons appear to be particularly vulnerable to areas of relative avascularity, referred to as critical zones. The vascular compromise of the supraspinatus is thought to be due to a number of factors: Although it is possible that sustained isometric contractions, prolonged adduction of the arm or increases in subacromial pressure64 may reduce the microcirculation, it is unlikely that frequent abduction or elevation of the arm would produce selective avascularity of the supraspinatus or biceps tendon. The close-packed position for the G-H joint is 90 degrees of G-H abduction and full ER or full abduction and ER, depending on the source. Without IR or ER occurring, the open-packed position of the G-H joint has traditionally been cited as 55 degrees of semi-abduction and 30 degrees of horizontal adduction.65 More recently, a cadaver study, which examined the point in the range at which maximal capsular laxity occurred in seven subjects, determined the open-packed position to be 39 degrees of abduction in the scapular plane or at the point which is 45% of the maximal available abduction range of motion (ROM).66 This finding suggests that the open-packed position may be closer to neutral and that joint mobility testing and joint mobilizations of the G-H joint should be initiated according to a smaller angle of abduction than the traditionally cited open-packed position.66 The zero position for the G-H joint is the arm relaxed by the side, which, relative to the scapula, averages approximately 0 degrees of abduction, 12 degrees of flexion, and 10 degrees of ER.67 According to Cyriax,68 the capsular pattern for the G-H joint is that ER is the most limited, abduction the next most limited, and IR the least limited in a 3:2:1 ratio, respectively. However, this pattern only appears to be consistent with adhesive capsulitis of the shoulder. IR, rather than ER or abduction, appears to be the most limited motion in conditions with selected capsular hypomobility.69 The A-C joint is a synovial joint, formed by the medial margin of the acromion and the lateral end of the clavicle. The A-C joint is most often described as a gliding or plane joint, but the joint surfaces can vary from flat to slightly convex or concave, corresponding with the articulating surface of the acromion. In the early stages of development, the articular surfaces of the A-C joint are lined with hyaline cartilage, which changes to fibrocartilage at approximately 17 years of age on the acromial side of the joint, and until approximately 24 years of age on the clavicular side.70 Within the A-C joint, there is variable presence of a thumbtack-shaped intra-articular fibrous disk that projects superiorly into, and incompletely divides, the A-C joint. This disk is subject to tearing.70 When viewed from above, the clavicle is convex anteriorly in the medial two-thirds and convex posteriorly in the lateral one-third (see Fig. 16-1). Variability in the inclination of the joint is common and can be anywhere from 10 to 50 degrees, but the anteromedial border of the acromion usually faces anteriorly, medially, and superiorly.71 This variability may account for the differences in vulnerability to separation seen among people.72 The A-C joint serves as the main articulation that suspends the upper extremity from the trunk, and it is at this joint about which the scapula moves. Specifically, the A-C joint provides the scapula with additional range of rotation on the thorax, which allows the scapula to adjust outside of its initial plane (posterior tipping and IR) to follow the changing shape of the thorax as movement occurs.73,74 The clavicle serves as the lever by which the upper extremity acts on the torso and as an attachment site for many soft tissues.70,75 The thin joint capsule is lined with synovium, which is strengthened anteriorly, posteriorly, inferiorly, and superiorly by A-C ligaments (see Fig. 16-6). The superior A-C ligament (see Fig. 16-6) gives support to the capsule and serves as the primary restraint to posterior translation and posterior axial rotation at the joint.70 Other support structures for the joint include the costoclavicular, coracoclavicular (conoid and trapezoid) ligament, the pectoralis major, SCM, deltoid, and trapezius muscles (see Fig. 16-10).75,76 FIGURE 16-10 Muscles of the shoulder. (Reproduced, with permission, from Chapter 1. Back. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) The coracoclavicular ligament (see Fig. 16-6), which is typically described as having conoid and trapezoid portions, is the primary support for the A-C joint and runs from the coracoid process to the inferior surface of the clavicle. The conoid ligament is the more fan shaped with its apex pointing inferiorly. It lies in the frontal plane and is the more medial of the two ligaments. This ligament functions to block coracoid movement away from the clavicle inferiorly.79 The trapezoid ligament arises from the medial border of the upper surface of the coracoid process and runs superiorly and laterally to insert into the inferior surface of the clavicle. It is larger, longer, and stronger than the conoid and forms a quadrilateral sheet that lies in a plane that is at right angles to the plane formed by the conoid ligament. The function of this ligament is unclear, although its orientation suggests that it may block medial movement of the coracoid,79 or act as a restraint to superior or posterior displacement of the clavicle.80 In addition, as the clavicle rotates upward, the coracoclavicular ligament dictates scapulothoracic rotation by virtue of its attachment to the scapula.81 Innervation to this joint is provided by the suprascapular, lateral pectoral, and axillary nerves (Fig. 16-8).83 The A-C joint receives its blood supply from branches of the suprascapular and thoracoacromial arteries (Fig. 16-9). Joints such as the A-C joint, which are not controlled by muscles, lack true capsular patterns. However, anecdotal clinical evidence suggests that the capsular pattern for the A-C joint is pain at the extremes of ROM, especially horizontal adduction and full elevation. The close-packed position for this joint seems to correspond to 90 degrees of G-H joint abduction. The open-packed position for this joint is undetermined, although it is likely to be when the arm is by the side. This positions the clavicle in approximately 15–20 degrees of retraction relative to the coronal plane and elevated approximately 2 degrees from the horizontal plane.84 The S-C joint, which is the only joint that connects the shoulder girdle to the axial skeleton, represents the articulation between the enlarged medial end of the clavicle, the clavicular notch of the manubrium of the sternum, and the cartilage of the first rib, which forms the floor of the joint. The articulating surfaces of the S-C joint are covered with fibrocartilage. The S-C joint is angulated slightly upward approximately 20 degrees in a posterior and lateral direction. If held vertically, the proximal end of the clavicle is convex whereas the manubrium surface is concave (see Fig. 16-1). If held anteroposteriorly, the proximal end of the clavicle is concave and the manubrium is convex. A very thick meniscus or disk is the key to the joint curvature. The clavicle presents with an irregularly shaped surface to the disk, and this lateral part of the joint acts as an ovoid. The disk is attached to the upper and posterior margin of the clavicle, and to the cartilage of the first rib by an intra-articular disk ligament, which functions to help prevent medial displacement of the clavicle, and completely divides the joint into two cavities: a larger one that is above and lateral to the disk, and a smaller one that is medial and below the disk. Greater movement occurs between the clavicle and the disk than between the disk and the manubrium. The S-C joint has very little bony stability and relies heavily on support from the surrounding capsule and ligaments. Some confusion seems to exist about classification of the S-C joint—it has been classified as a ball and socket joint,83 a plane joint, and as a saddle joint.83 These classifications depend on whether anatomy or function is being considered. For example, when its function is considered, the S-C joint acts similar to a ball and socket joint, allowing for motion in almost all planes, including rotation. Because of its bony arrangement, the S-C joint itself is extremely weak, but is held securely by strong ligaments that hold the sternal end of the clavicle downward and toward the sternum. These ligaments include the following: Capsular Ligament. This ligament represents the inner aspect of the joint capsule. The anterior portion is heavier and stronger, and it covers the anterior aspect of the joint running obliquely from the proximal end of the clavicle to the sternum in an inferior and medial direction. The posterior component, which covers the posterior aspect of the joint, also runs obliquely from the proximal end of the clavicle to the sternum in an inferior and medial direction. The capsular ligament is considered the most important and strongest ligaments of the S-C joint and is most responsible for preventing upward displacement of the medial clavicle in the presence of a downward force on the distal end of the shoulder.86 Interclavicular. This ligament connects the superomedial sternal ends of each clavicle with the capsular ligaments and the upper sternum. Together, these structures help to hold up the shoulder when the upper extremities are resting by the side. This function can be demonstrated by placing a finger in the superior sternal notch; the ligament is lax with elevation of the arm but becomes taut when both arms are hanging by the sides. Costoclavicular (Rhomboid Ligament). This short and strong ligament, which runs from the upper border of the first rib to the inferior surface of the clavicle, consists of two laminae: fibers of the anterior lamina run superiorly and laterally and check elevation and lateral movement (upward rotation) of the clavicle.86 The posterior lamina fibers run superiorly and medially and check downward rotation of the clavicle.86 Due to the support provided by the ligaments, the S-C joint is very stable and trauma to the clavicle usually results in a fracture rather than a joint dislocation.87 Gross motions occur here as with the A-C joint. The SCM, which can be seen clearly with rotation of the head, has a tendinous sternal and clavicular insertion. The subclavius (C5–6) has a questionable function but may function as a dynamic ligament, which contracts and pulls the clavicle toward the manubrium. The close-packed position for the S-C joint is maximum arm elevation and protraction. The open-packed position for the S-C joint has yet to be determined, but is likely to be when the arm is by the side. Similar to the A-C joint, the S-C joint is not controlled by muscles and therefore lacks a specific capsular pattern. One possibility, seen clinically, is pain at the extreme ranges of motion, especially full arm elevation and horizontal adduction. Pain can be referred from this joint to the throat, anterior chest, and axillae. The neural supply to this joint is primarily from the following: The S-C joint receives its blood supply from the internal thoracic and suprascapular arteries.88 This articulation is functionally a joint, but it lacks the anatomic characteristics of a true synovial joint. However, the movement of the scapula on the wall of the thoracic cage is critical to shoulder joint motion. A lack of ligamentous support at this “joint” delegates the function of stability fully to the muscles that attach the scapula to the thorax. An altered position of the scapula, or an abnormal motion at this joint due to muscle imbalances, have both been linked with shoulder complex dysfunction.24,89,90 Motions at this joint are described according to the movement of the scapula relative to the thorax. Available motion consists of approximately 60 degrees of upward rotation of the scapula, 40–60 degrees of IR/ER, and 30–40 degrees of anterior and posterior tipping of the scapula.91 Other motions occurring here include elevation and depression and retraction (adduction) and protraction (abduction) of the scapula (Fig. 16-11). FIGURE 16-11 Movements of the scapula and upper extremity. (Reproduced, with permission, from Chapter 30. Shoulder and Axilla. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) Since the scapulothoracic joint is not a true joint, it is difficult to determine its close-packed position or capsular pattern. Relative to the thorax, when the arm is by the side, the scapula is in an average of 30–45 degrees of IR, and slight upward rotation, and approximately 5–20 degrees of anterior tipping.67,91 There are a number of bursae located in and around the scapulothoracic articulation. The scapulothoracic bursa is located between the thoracic cage and the deep surface of the serratus anterior.92 The subscapular bursa is most often located between the superficial surface of the serratus anterior and the subscapularis.92 The scapulotrapezial bursa lies between the middle and lower trapezius fibers and the superomedial scapula.92 The purpose and clinical significance of the scapulotrapezial bursa are not known. It may encourage smooth gliding of the superomedial angle of the scapula against the undersurface of the trapezius during scapular rotation in the same manner that the scapulothoracic bursa (between the serratus attachment at the anteromedial surface of the superior angle) encourages smooth gliding against the underlying ribs.92 It is possible that inflammation of either of these bursae, directly or as a consequence of injury, may result in painful clicking at the superomedial angle of the scapula.92 The relationship of the spinal accessory nerve to the scapulotrapezial bursa may also have clinical importance, especially as it is closely applied to the superficial wall of the scapulotrapezial bursa.92 The spinal accessory nerve receives afferent fibers from C3 and C4, which are thought to be proprioceptive, before reaching the deep surface of the trapezius.92,93 As a consequence of their proximity, inflammation and fibrosis within the bursa may cause irritation and pain of the nerve, or interference with the normal proprioceptive feedback mechanism provided by the nerve.92 A number of significant muscles control motion at the shoulder and provide dynamic stabilization. Rarely does a single muscle act in isolation at the shoulder. For simplicity, the muscles acting at the shoulder may be described in terms of their functional roles: scapular pivoters, humeral propellers, humeral positioners, and shoulder protectors (Table 16-2).94 TABLE 16-2 Muscles of the Shoulder Complex Scapular pivoters Trapezius Serratus anterior Levator scapulae Rhomboid major Rhomboid minor Humeral propellers Latissimus dorsi Teres major Pectoralis major Pectoralis minor Humeral positioners Deltoid Shoulder protectors Rotator cuff (supraspinatus, infraspinatus, teres minor, and subscapularis) Long head of the biceps brachii The scapular pivoters comprise the trapezius, serratus anterior, levator scapulae, rhomboid major, and rhomboid minor.94 As a group, these muscles are involved with motions at the scapulothoracic articulation, and their proper function is vital to the normal biomechanics of the whole shoulder complex. The scapular muscles can contract isometrically, concentrically, or eccentrically, depending on the desired action and whether the action involves stabilization, acceleration, or deceleration. To varying degrees, the serratus anterior and all parts of the trapezius cooperate during the upward rotation of the scapula. The trapezius muscle (Fig. 16-10) originates from the medial third of the superior nuchal line, the external occipital protuberance, the ligamentum nuchae, the apices of the seventh cervical vertebra, all the thoracic spinous processes, and the supraspinous ligaments of the cervical and thoracic vertebrae. The upper fibers descend to attach to the lateral third of the posterior border of the clavicle. The middle fibers of the trapezius run horizontally to the medial acromial margin and superior lip of the spine of the scapula. The inferior fibers ascend to attach to an aponeurosis gliding over a smooth triangular surface at the medial end of the spine of the scapula to a tubercle at the scapular lateral apex. It has been suggested that the upper fibers of this muscle have a different motor supply than that to the middle and lower portions.95,96 Recent clinical and anatomical evidence seems to suggest that the spinal accessory nerve provides the most important and consistent motor supply to all portions of the trapezius muscle, and although the C2–4 branches of the cervical plexus are present, no particular elements of innervation within the trapezius have been determined.97 One of the functions of the trapezius is to produce shoulder girdle elevation on a fixed cervical spine. For the trapezius to perform its actions, the cervical spine must first be stabilized by the anterior neck flexors to prevent simultaneous occipital extension from occurring. Failure to prevent this occipital extension would allow the head to translate anteriorly, resulting in a decrease in the length, and therefore the efficiency, of the trapezius,98 and an increase in the cervical lordosis. The muscular digitations of the serratus anterior (see Fig. 16-5) originate from the upper eight to ten ribs and fascia over the intercostals. The muscle is composed of three functional components:100,101 The serratus anterior is activated with all shoulder movements, but especially during shoulder flexion and abduction.101 Working in synergy with the trapezius, as part of a force couple (see later), the main function of the serratus anterior is to protract and upwardly rotate the scapula,102,103 while providing a strong, mobile base of support to position the glenoid optimally for maximum efficiency of the upper extremity.104 Its lower fibers draw the lower angle of the scapula forward to rotate the scapula upward while maintaining the scapula on the thorax during arm elevation.105 This moves the coracoacromial arch out of the path of the advancing greater tuberosity and opposes the excessive elevation of the scapula by the levator scapulae and trapezius muscles.106 Without upward rotation and protraction of the scapula by the serratus anterior, full G-H elevation is not possible. In fact, in patients with complete paralysis of the serratus anterior, Gregg et al.104 reported that abduction is limited to 110 degrees. Dysfunction of the serratus anterior muscle causes winging of the scapula as the patient attempts to elevate the arm.4,107 Scapulothoracic dysfunction can also contribute to G-H instability, as the normal stable base of the scapula is destabilized during abduction or flexion.4,108,109 The serratus anterior muscle is innervated by the long thoracic nerve (C5–7). The levator scapulae muscle (see Fig. 16-10) originates by tendinous strips from the transverse processes of the atlas, axis, and C3 and C4 vertebrae, and descends diagonally to insert into the medial superior angle of the scapula. The levator scapulae can act on the cervical spine (see Chapter 23) and on the scapula. If it acts on the cervical spine, it can produce extension, side flexion, and rotation of the cervical spine to the same side.110 When acting on the scapula during upper extremity flexion or abduction, the levator scapula muscle acts as an antagonist to the trapezius muscle, and provides eccentric control of scapular upward rotation in the higher ranges of motion (see later).111 Both the trapezius and levator scapulae muscles are activated with increased upper extremity loads.98,101,112 The levator scapulae muscle is innervated by the posterior (dorsal) scapular nerve (C3–5). The rhomboid major muscle (Fig. 16-10) originates from the second to fifth thoracic spinous processes and the overlying supraspinous ligaments. The fibers descend to insert into the medial scapular border between the root of the scapular spine and the inferior angle of the scapula. The rhomboid minor muscle (see Fig. 16-10) originates from the lower aspect of the ligamentum nuchae, and the seventh cervical and first thoracic spinous processes, and attaches to the medial border of the scapula at the root of the spine of the scapula. The rhomboid muscles help control scapular positioning, particularly with horizontal flexion and extension of the shoulder complex.111 The rhomboid muscles are innervated by the posterior (dorsal) scapular nerve (C4–5). The total muscle mass of the shoulder’s internal rotators (subscapularis, anterior deltoid, pectoralis major, latissimus dorsi, and teres major) is much greater than that of the external rotators (infraspinatus, teres minor, and posterior deltoid).99 This fact explains why the shoulder internal rotators produce approximately 1.75 times greater isometric torque than the external rotators.113 Peak torques of the internal rotators also exceed the external rotators when measured isokinetically, under both concentric and eccentric conditions.99,114 The latissimus dorsi muscle (see Fig. 16-12) originates from the spinous processes of the last six thoracic vertebrae, the lower three or four ribs, the lumbar and sacral spinous processes through the thoracolumbar fascia, the posterior third of the external lip of the iliac crest, and a slip from the inferior scapular angle. The scapular slip allows the latissimus dorsi to act at the scapulothoracic articulation. The latissimus dorsi inserts into the intertubercular sulcus of the humerus. The muscle functions as an extensor, adductor, and powerful internal rotator of the humerus, and also assists in scapular depression, retraction, and downward rotation.115 It is innervated by the thoracodorsal nerve (C6–8). FIGURE 16-12 Latissimus dorsi. (Reproduced, with permission, from Chapter 30. Shoulder and Axilla. In: Morton DA, Foreman K, Albertine KH. eds. The Big Picture: Gross Anatomy. New York, NY: McGraw-Hill; 2011.) The teres major (see Fig. 16-12) originates from the inferior third of the lateral border of the scapula and just superior to the inferior angle. The teres major tendon inserts into the medial lip of the intertubercular groove of the humerus. The teres major functions to complement the actions of the latissimus dorsi in that it extends, adducts, and internally rotates the G-H joint. It is innervated by the lower subscapular nerve (C5, C6). The pectoralis major (see Fig. 16-13) originates from the sternal half of the clavicle, half of the anterior surface of the sternum to the level of the sixth or seventh costal cartilage, the sternal end of the sixth rib, and the aponeurosis of the obliquus externus abdominis. The fibers of the pectoralis major converge to form a tendon that inserts into the lateral lip of the intertubercular sulcus of the humerus. Although this muscle does not insert into the scapula, it does act upon the scapulothoracic articulation through its insertion on the humerus. The function of the pectoralis muscle depends on which fibers are activated: FIGURE 16-13 Pectoralis major muscle. The pectoralis minor (see Fig. 16-9) originates from the outer surface of the upper margins of the third to fifth ribs near their cartilage. The fibers of the pectoralis minor ascend laterally, converging to a tendon that inserts into the coracoid process of the scapula. The pectoralis minor muscle is innervated by the medial pectoral nerve (C6–8). The deltoid muscle originates from the lateral third of the clavicle, the superior surface of the acromion, and the spine of the scapula (Fig. 16-13). It inserts into the deltoid tuberosity of the humerus. The deltoid can be described as three separate muscles—anterior, middle, and posterior—all of which function as humeral positioners, positioning the humerus in space.94 The deltoid muscle is innervated by the axillary nerve (C5–6). The rotator cuff muscles (Fig. 16-4), which consist of the supraspinatus, infraspinatus, teres minor, and subscapularis, are commonly implicated with shoulder pathology. The anatomy of these muscles was described previously (see “Glenohumeral Joint” section). These muscles are referred to as the protectors of the shoulder since, in addition to actively moving the humerus, they fine-tune the humeral head position during arm elevation.94 Compared with most joints that have a single axis on which torques are generated, the shoulder is very different, because it has no fixed axis. As a result, each muscle activation creates a unique set of rotational moments, which necessitates precise coordination in the timing and magnitude of muscle contractions.72 Jenp et al.116 used electromyography (EMG) to detect the most specific positions of the highest activation for the individual rotator cuff muscles. The greatest activation of the subscapularis was with the arm in the scapular plane at 90 degrees of elevation and neutral humeral rotation. The subscapularis has also been shown to be an effective humeral head depressor in ER, whereas it produces almost no A-P translation in abduction and ER. The infraspinatus–teres minor muscles are very effective humeral head depressors with the arm in the sagittal plane and the humerus elevated to 90 degrees in the midrange of external/IR (the so-called “hornblower’s” position). The supraspinatus has yet to be effectively isolated. The rotator cuff muscles have an important role in the function of the shoulder and serve the following:
CHAPTER 16
The Shoulder
OVERVIEW
 Suspending the upper limb.
Suspending the upper limb.
 Providing sufficient fixation so that motion of the upper extremity or trunk can occur.
Providing sufficient fixation so that motion of the upper extremity or trunk can occur.
 Serving as a fulcrum for arm elevation. Three types of arm elevation are recognized: an upward motion of the upper extremity in the scapular plane (scaption) and the motions in either the coronal plane (abduction) or in the sagittal plane (flexion).
Serving as a fulcrum for arm elevation. Three types of arm elevation are recognized: an upward motion of the upper extremity in the scapular plane (scaption) and the motions in either the coronal plane (abduction) or in the sagittal plane (flexion).
ANATOMY
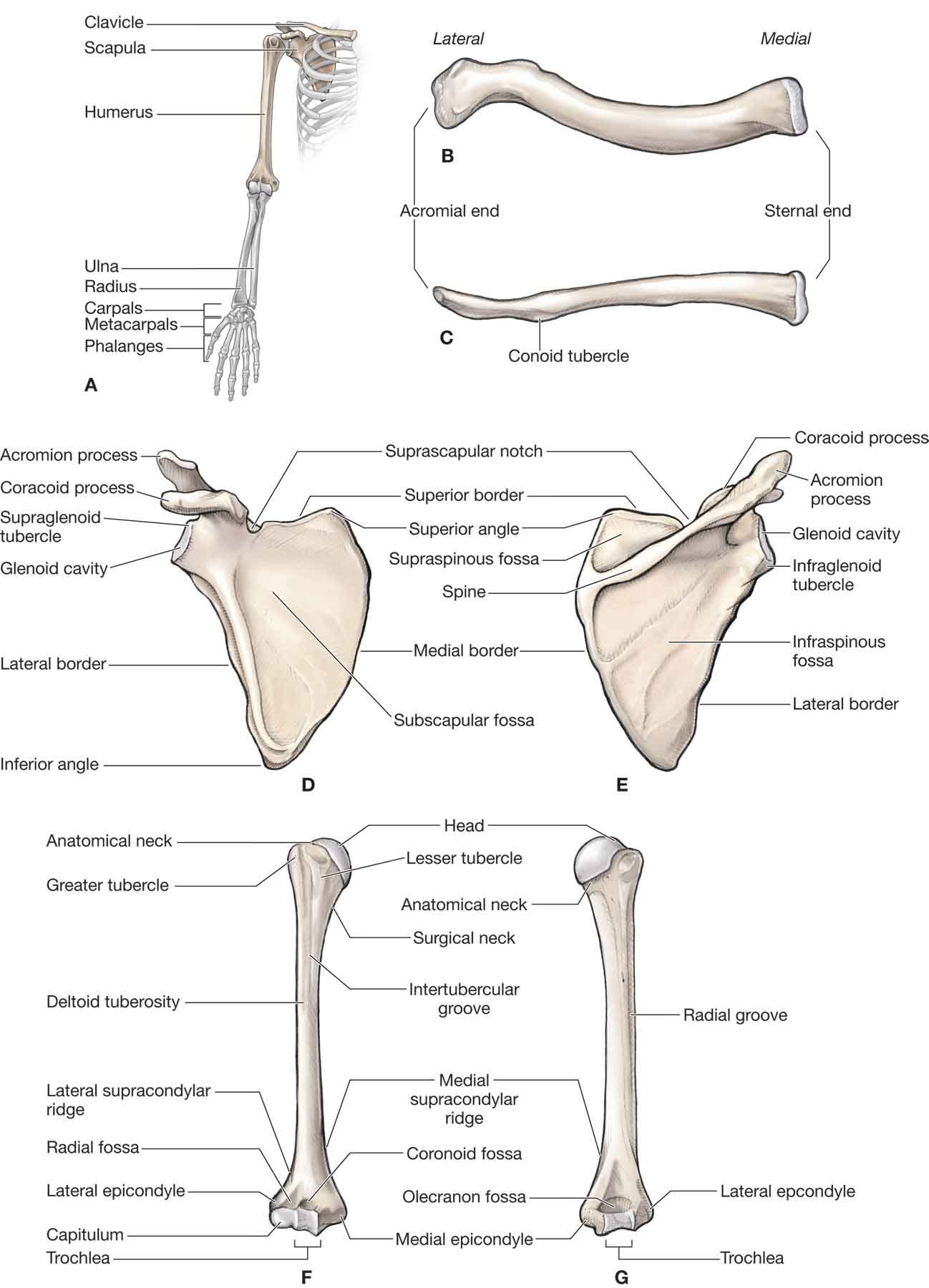
 three bones (the humerus, the clavicle, and the scapula);
three bones (the humerus, the clavicle, and the scapula);
 three joints (the sternoclavicular [S-C], the acromioclavicular [A-C], and the glenohumeral [G-H] joints);
three joints (the sternoclavicular [S-C], the acromioclavicular [A-C], and the glenohumeral [G-H] joints);
 one “pseudojoint” (the articulation between the scapula and the thorax);
one “pseudojoint” (the articulation between the scapula and the thorax);
 one physiological area (the suprahumeral or subacromial space).
one physiological area (the suprahumeral or subacromial space).
GLENOHUMERAL JOINT
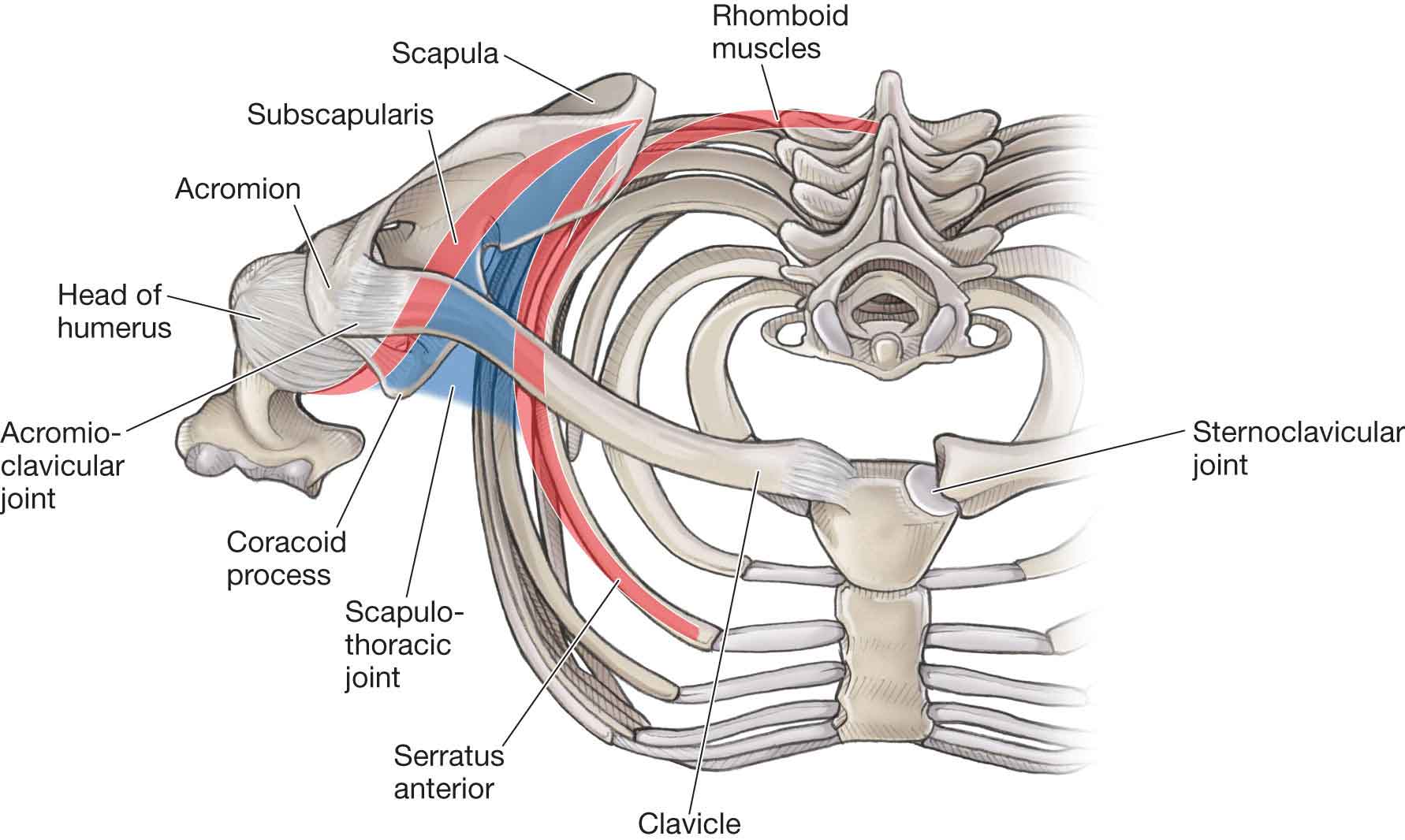
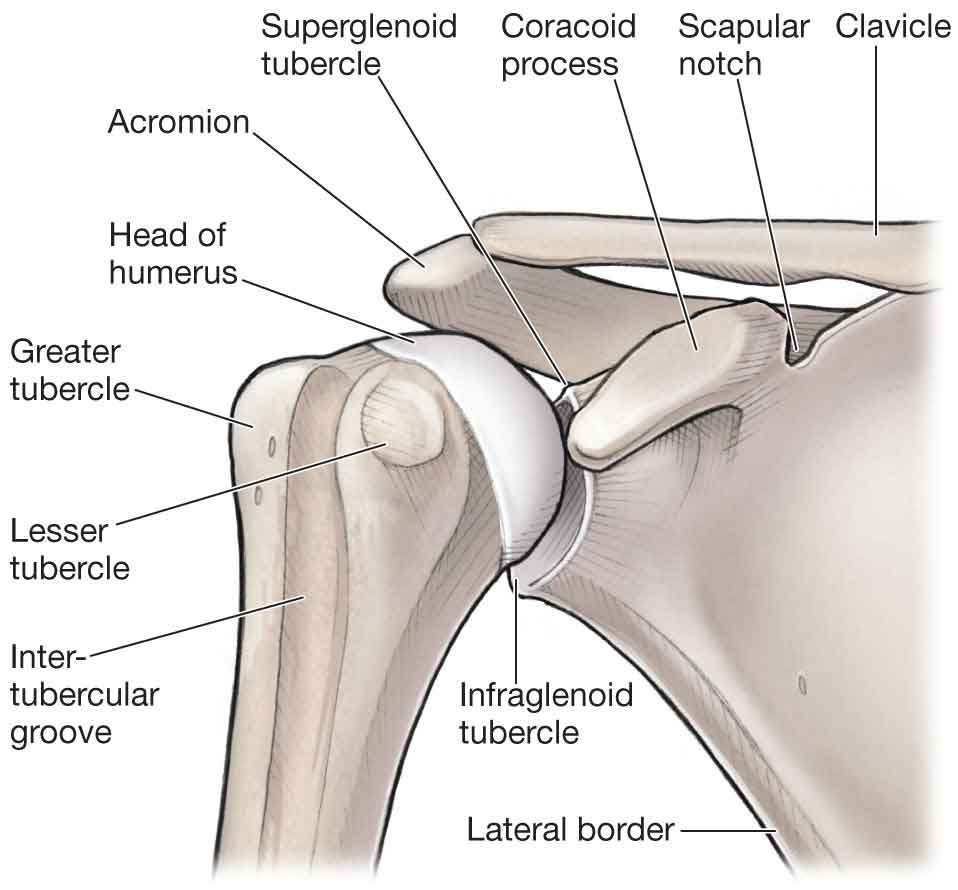
 adduction, flexion, and internal rotation (IR);
adduction, flexion, and internal rotation (IR);
 abduction and elevation;
abduction and elevation;
 adducted at the side, with the scapula rotated downward.
adducted at the side, with the scapula rotated downward.
 Type I has a relatively flat undersurface
Type I has a relatively flat undersurface
 Type II is slightly convex
Type II is slightly convex
 Type III is hooked
Type III is hooked
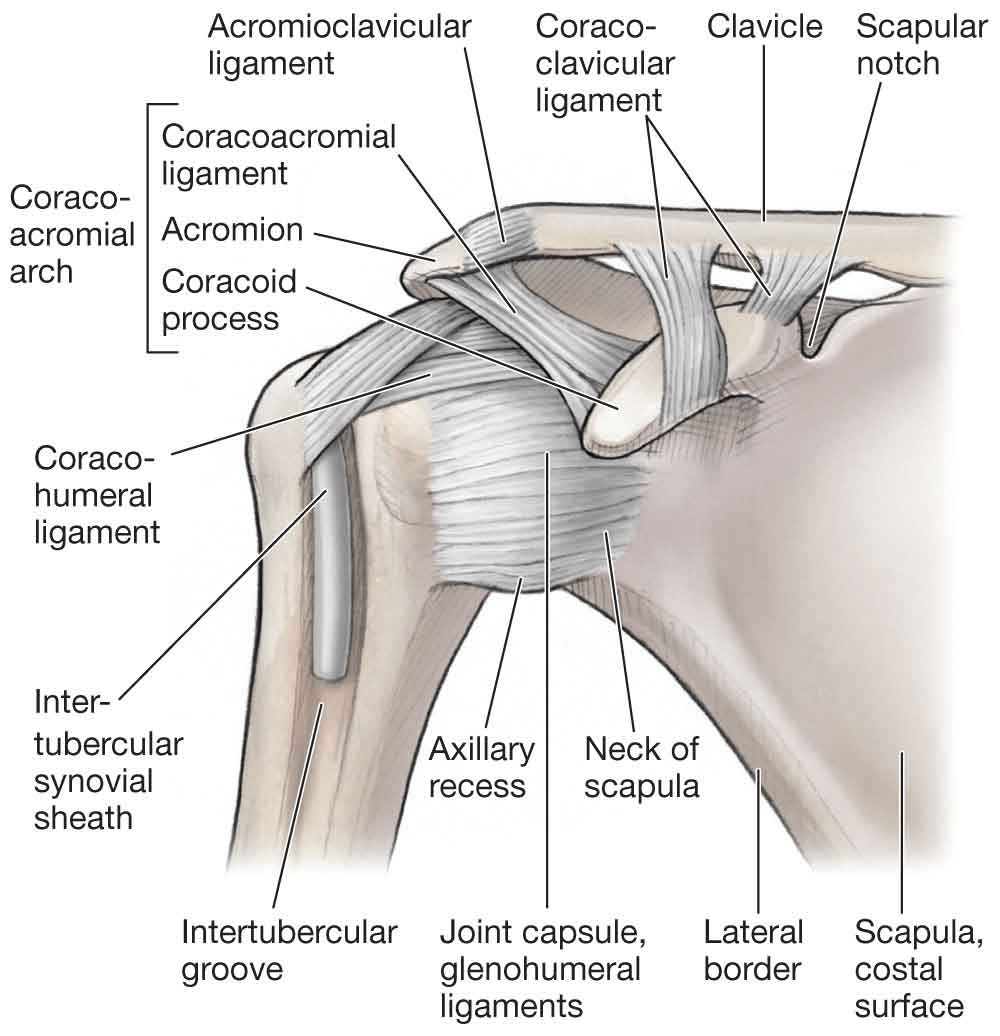
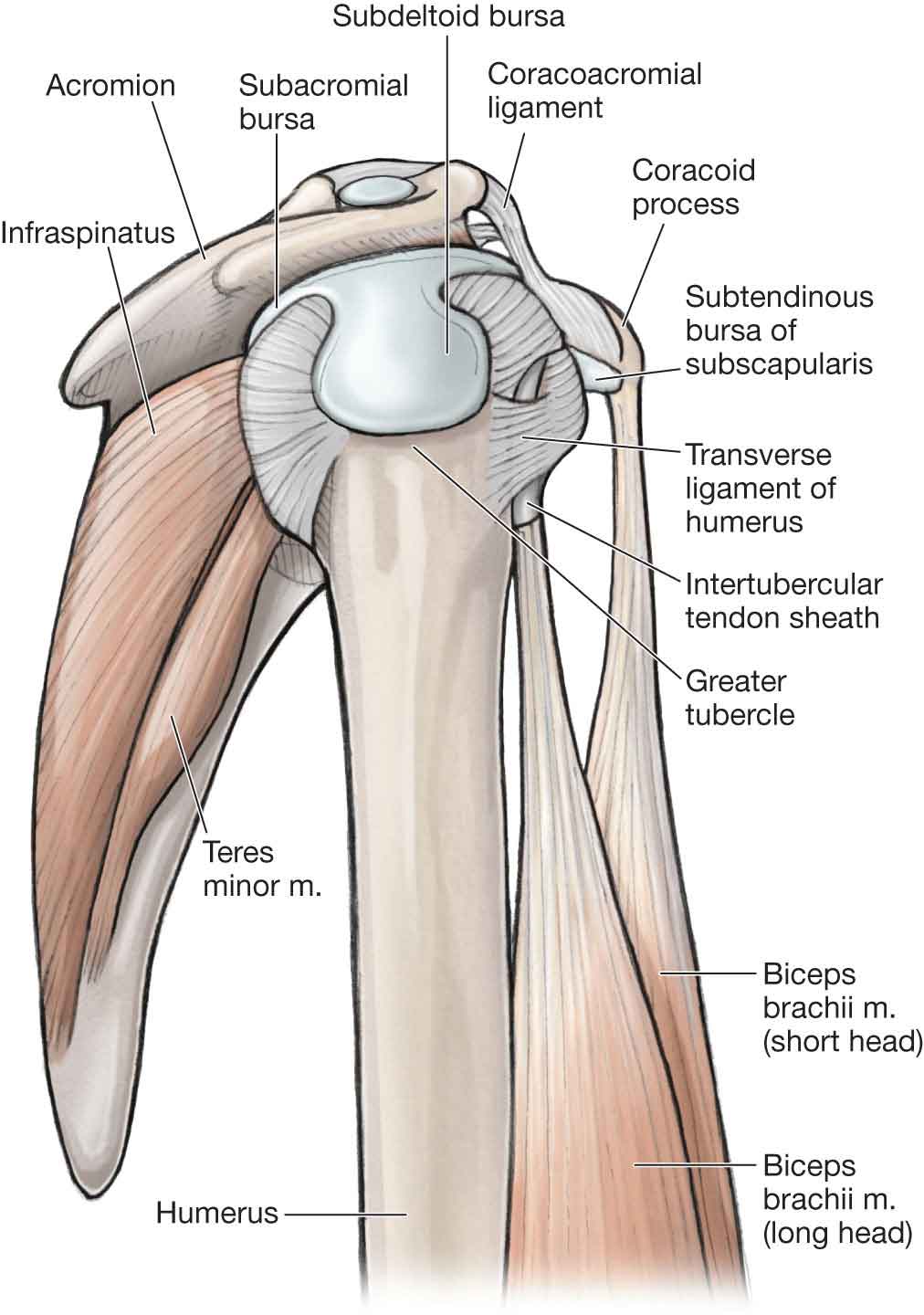
Ligaments
 The anterior G-H ligament is under tension when the shoulder is in extension, abduction, and/or external rotation (ER).
The anterior G-H ligament is under tension when the shoulder is in extension, abduction, and/or external rotation (ER).
 The posterior G-H ligament is under tension in flexion and ER.
The posterior G-H ligament is under tension in flexion and ER.
 The inferior G-H ligament is the most important of the G-H ligaments. It is under tension when the shoulder is abducted, extended, and/or externally rotated. In addition, this ligament is a primary restraint against both anterior and posterior dislocations of the humeral head, and is the most important stabilizing structure of the shoulder in the overhead athlete.34 At 0-degree abduction, the subscapularis muscle, the labrum, and the superior G-H ligament are the primary restraints against anterior translation. At 45-degree abduction, the subscapularis muscle and the middle and inferior G-H ligaments, along with labrum, prevent anterior translation. When the arm is abducted more than 90 degrees, which is the most common position of anterior dislocation, the anterior fibers of the inferior G-H ligament are the primary restraint against anterior movement.9,35
The inferior G-H ligament is the most important of the G-H ligaments. It is under tension when the shoulder is abducted, extended, and/or externally rotated. In addition, this ligament is a primary restraint against both anterior and posterior dislocations of the humeral head, and is the most important stabilizing structure of the shoulder in the overhead athlete.34 At 0-degree abduction, the subscapularis muscle, the labrum, and the superior G-H ligament are the primary restraints against anterior translation. At 45-degree abduction, the subscapularis muscle and the middle and inferior G-H ligaments, along with labrum, prevent anterior translation. When the arm is abducted more than 90 degrees, which is the most common position of anterior dislocation, the anterior fibers of the inferior G-H ligament are the primary restraint against anterior movement.9,35
 The middle G-H ligament is under tension when the shoulder is flexed and externally rotated. In addition, the middle G-H ligament and the subscapularis tendon limit ER from 45–75 degrees of abduction, and are important anterior stabilizers.
The middle G-H ligament is under tension when the shoulder is flexed and externally rotated. In addition, the middle G-H ligament and the subscapularis tendon limit ER from 45–75 degrees of abduction, and are important anterior stabilizers.
 The coracohumeral ligament. The coracohumeral ligament (see Fig. 16-6) arises from the lateral end of the coracoid process and runs laterally, where it is split into two bands by the presence of the biceps tendon. The posterior band blends with the supraspinatus tendon to insert near the greater tuberosity, and the anterior band blends with the subscapularis tendon to insert near the lesser tuberosity. The coracohumeral ligament covers the superior G-H ligament anterosuperiorly and fills the space between the tendons of the supraspinatus and subscapularis muscles, uniting these tendons to complete the rotator cuff in this area. Tears of the cuff usually extend longitudinally between the supraspinatus and coracohumeral ligament so that the hood action of the cuff is lost. It is generally agreed that the posterior band of the coracohumeral ligament limits flexion, whereas the anterior band limits extension of the G-H joint.36 Both the bands also limit inferior and posterior translation of the humeral head, strengthening the superoanterior aspect of the capsule.36,37
The coracohumeral ligament. The coracohumeral ligament (see Fig. 16-6) arises from the lateral end of the coracoid process and runs laterally, where it is split into two bands by the presence of the biceps tendon. The posterior band blends with the supraspinatus tendon to insert near the greater tuberosity, and the anterior band blends with the subscapularis tendon to insert near the lesser tuberosity. The coracohumeral ligament covers the superior G-H ligament anterosuperiorly and fills the space between the tendons of the supraspinatus and subscapularis muscles, uniting these tendons to complete the rotator cuff in this area. Tears of the cuff usually extend longitudinally between the supraspinatus and coracohumeral ligament so that the hood action of the cuff is lost. It is generally agreed that the posterior band of the coracohumeral ligament limits flexion, whereas the anterior band limits extension of the G-H joint.36 Both the bands also limit inferior and posterior translation of the humeral head, strengthening the superoanterior aspect of the capsule.36,37
 The coracoacromial ligament. The coracoacromial ligament (see Fig. 16-6) is often described as the roof of the shoulder. It is a very thick structure that runs from the coracoid process to the anteroinferior aspect of the acromion, with some of its fibers extending to the A-C joint. The ligament consists of two bands that join near the acromion, and it is ideally suited, both anatomically and morphologically, to prevent separation of the A-C joint surfaces. The coracoclavicular ligaments and the costoclavicular ligaments are described in the A-C joint section and the S-C joint section, respectively.
The coracoacromial ligament. The coracoacromial ligament (see Fig. 16-6) is often described as the roof of the shoulder. It is a very thick structure that runs from the coracoid process to the anteroinferior aspect of the acromion, with some of its fibers extending to the A-C joint. The ligament consists of two bands that join near the acromion, and it is ideally suited, both anatomically and morphologically, to prevent separation of the A-C joint surfaces. The coracoclavicular ligaments and the costoclavicular ligaments are described in the A-C joint section and the S-C joint section, respectively.
Coracoacromial Arch
 the head of the humerus,
the head of the humerus,
 the long head of biceps tendon (intra-articular portion),
the long head of biceps tendon (intra-articular portion),
 the superior aspect of the joint capsule,
the superior aspect of the joint capsule,
 the supraspinatus and upper margins of subscapularis and infraspinatus,
the supraspinatus and upper margins of subscapularis and infraspinatus,
 the subdeltoid–subacromial bursae,
the subdeltoid–subacromial bursae,
 the inferior surface of coracoacromial arch.
the inferior surface of coracoacromial arch.
Bursae
 The subcoracoid bursa. This bursa is located under the coracoid process.
The subcoracoid bursa. This bursa is located under the coracoid process.
 The subscapular bursa. This bursa is located between the subscapular muscle tendon and the anterior neck of the scapula and protects the tendon as it passes under the coracoid process.
The subscapular bursa. This bursa is located between the subscapular muscle tendon and the anterior neck of the scapula and protects the tendon as it passes under the coracoid process.
Neurology
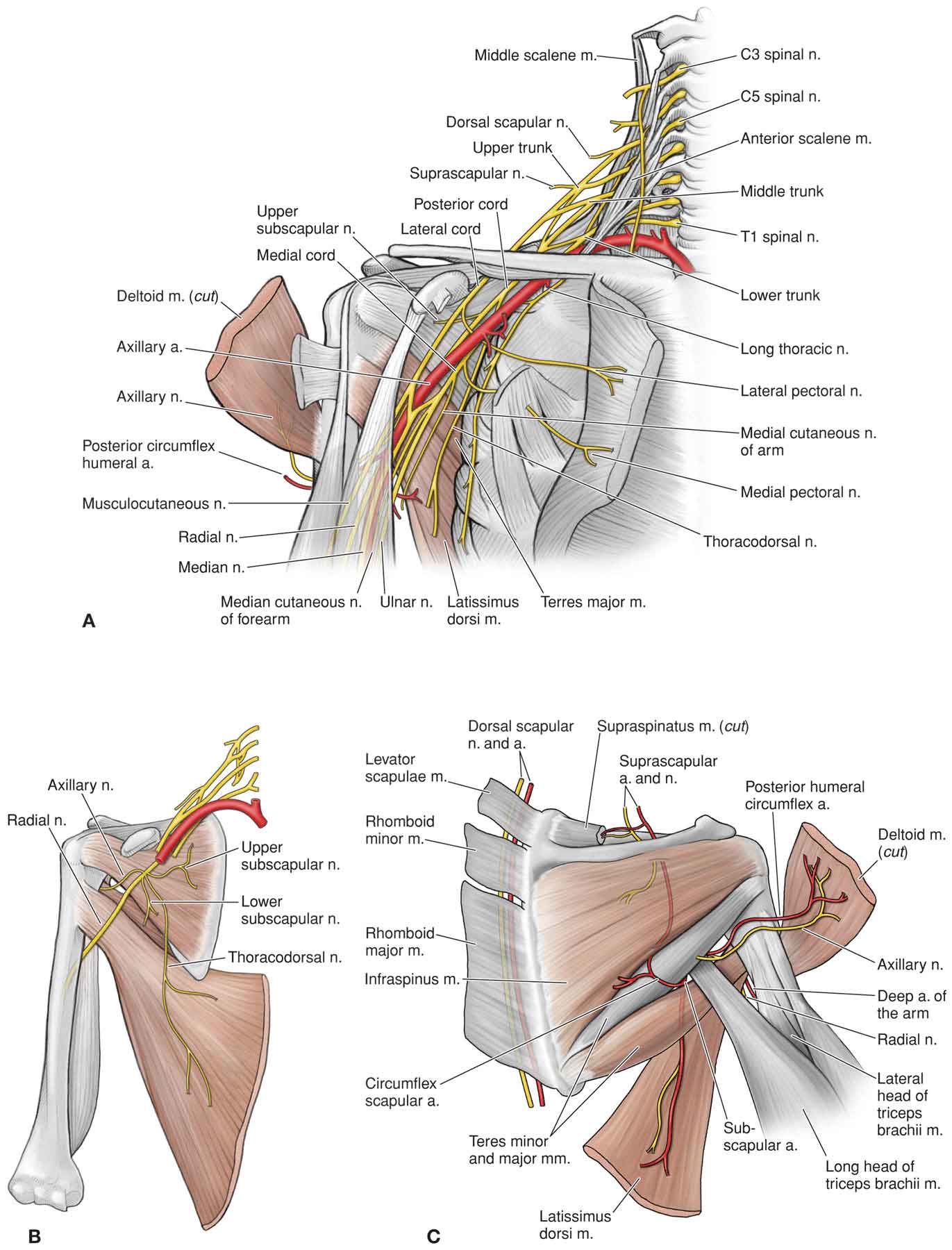
Vascularization
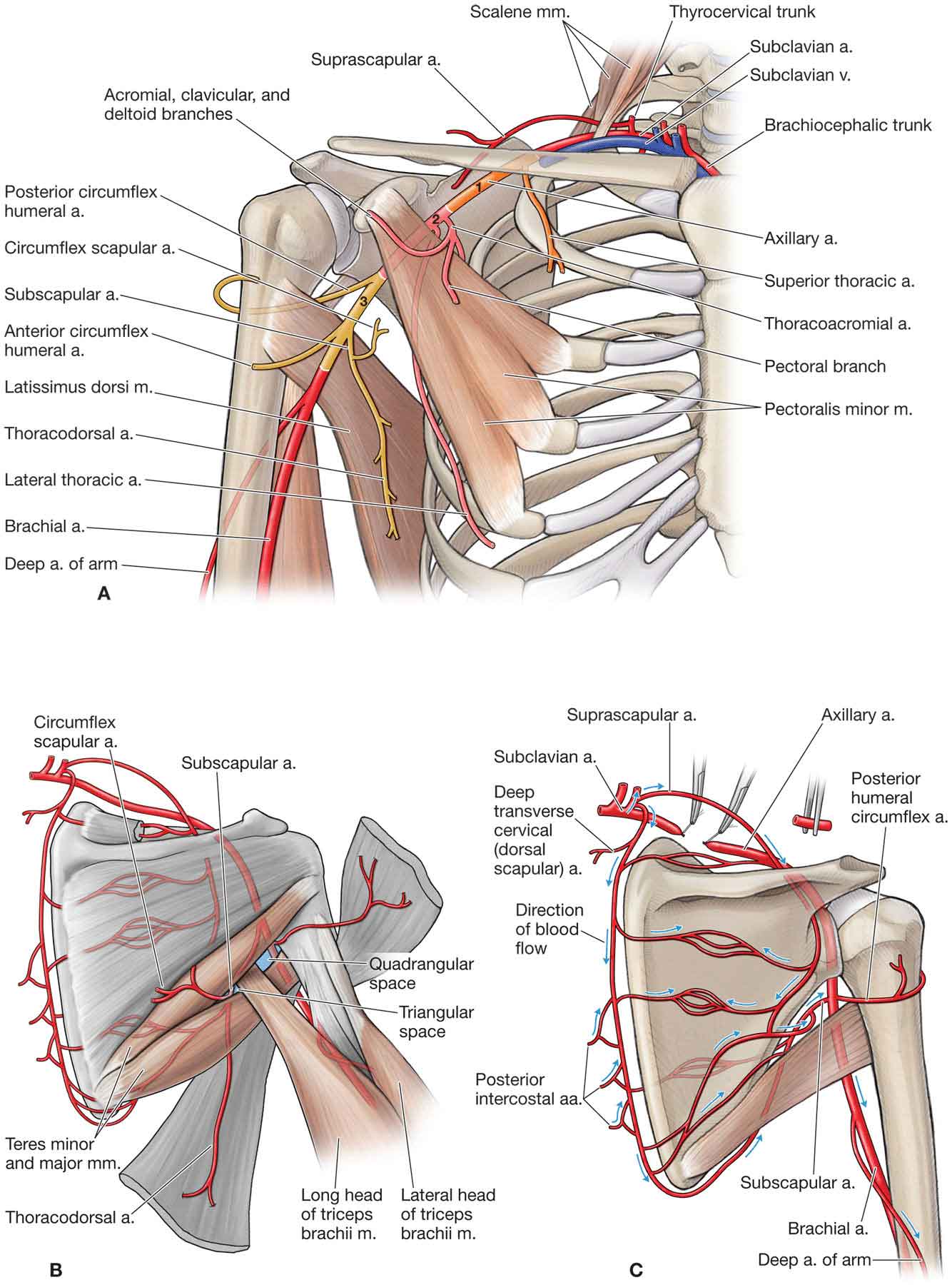
 It can be directly compressed by the subacromial structures. With the arm adducted to the side, the vessels within the supraspinatus tendon are poorly perfused.56 Other arm positions, such as raising the arm above 30 degrees, have been shown to increase intramuscular pressure in the supraspinatus muscle to an extent that may impair normal blood perfusion.56,57
It can be directly compressed by the subacromial structures. With the arm adducted to the side, the vessels within the supraspinatus tendon are poorly perfused.56 Other arm positions, such as raising the arm above 30 degrees, have been shown to increase intramuscular pressure in the supraspinatus muscle to an extent that may impair normal blood perfusion.56,57
 Its blood vessels travel parallel to the tendon fibers, which make them vulnerable to stretch.58 Avascularity appears to increase with age beginning as early as 20 years.59
Its blood vessels travel parallel to the tendon fibers, which make them vulnerable to stretch.58 Avascularity appears to increase with age beginning as early as 20 years.59
 The presence of a critical zone just proximal to the supraspinatus insertion point.56 Two early studies noted a critical zone that lies slightly proximal to the supraspinatus insertion point.60,61 Since then, it has been determined that the critical zone is more likely a zone of anastomoses between the vessels supplying the bone and the tendon and is not less vascular except in certain positions.56,62,63
The presence of a critical zone just proximal to the supraspinatus insertion point.56 Two early studies noted a critical zone that lies slightly proximal to the supraspinatus insertion point.60,61 Since then, it has been determined that the critical zone is more likely a zone of anastomoses between the vessels supplying the bone and the tendon and is not less vascular except in certain positions.56,62,63
Close-Packed Position
Open-Packed Position
Capsular Pattern
THE ACROMIOCLAVICULAR JOINT
Ligaments
Neurology
Vascularization
Capsular Pattern
Close-Packed Position
Open-Packed Position
STERNOCLAVICULAR JOINT
Ligaments
Capsular-Ligamentous Structures
Close-Packed Position
Open-Packed Position
Capsular Pattern
Neurology
 The anterior supraclavicular nerve
The anterior supraclavicular nerve
 The nerve to the subclavius (medial accessory phrenic) C5–6
The nerve to the subclavius (medial accessory phrenic) C5–6
 The T1 spinal nerve root
The T1 spinal nerve root
Vascularization
SCAPULOTHORACIC JOINT
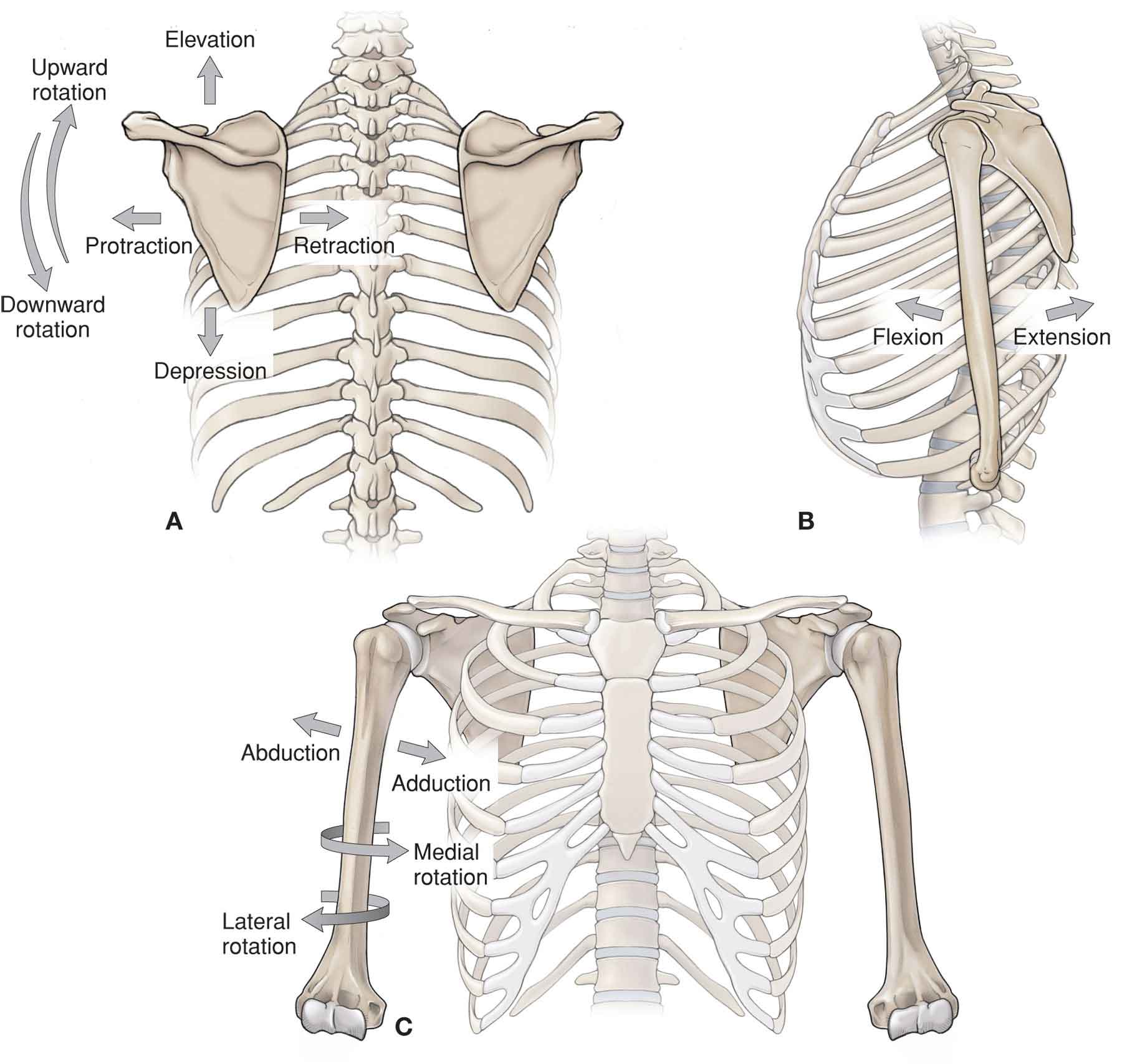
Close-Packed and Capsular Pattern
Open-Packed Position
Bursae
MUSCLES OF THE SHOULDER COMPLEX
Scapular Pivoters
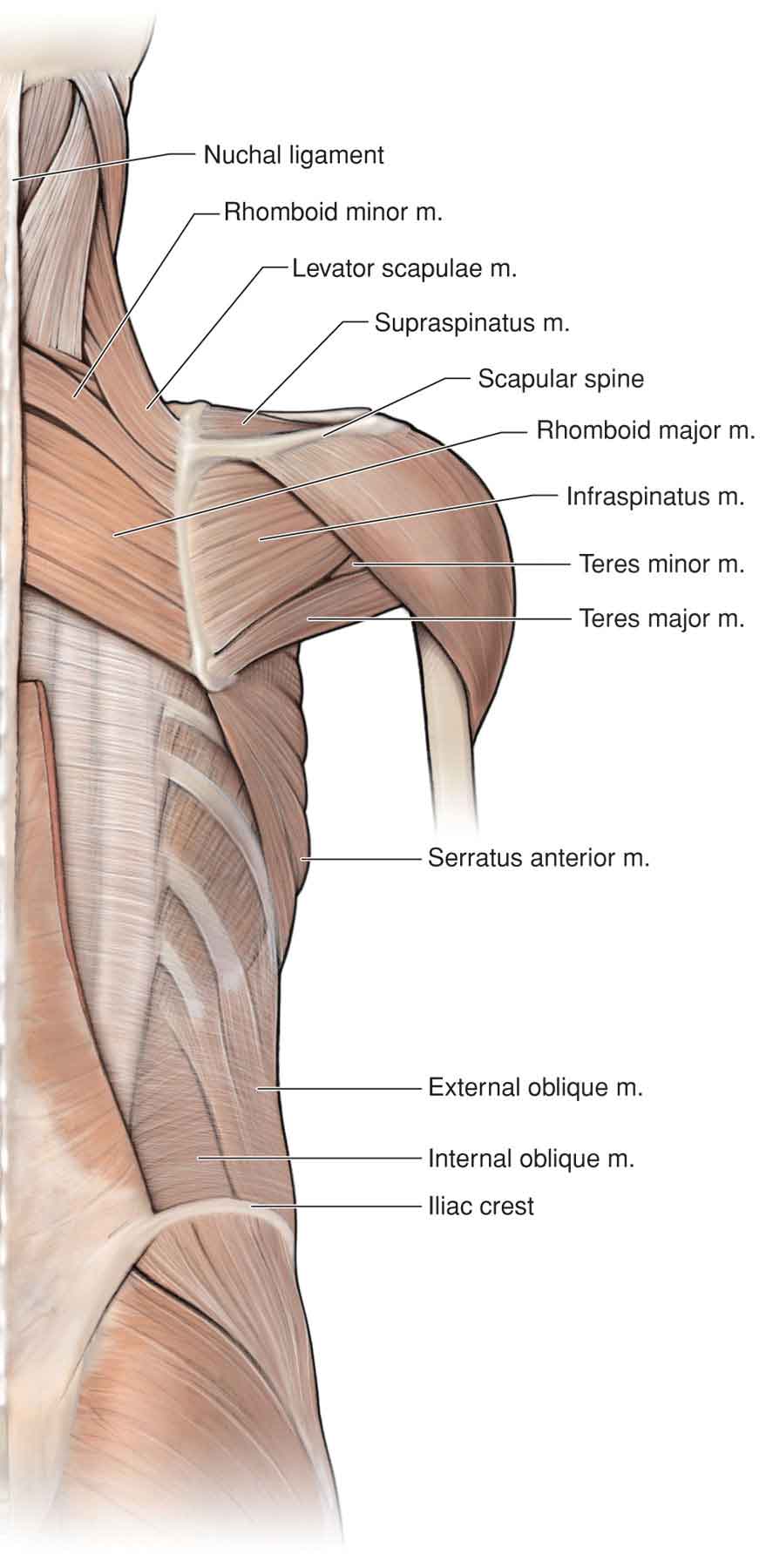
Trapezius
Serratus Anterior
 The upper component originates from the first and second ribs and inserts into the superior angle of the scapula.
The upper component originates from the first and second ribs and inserts into the superior angle of the scapula.
 The middle component arises from the second, third, and fourth ribs and inserts into the anterior aspect of the medial scapular border.
The middle component arises from the second, third, and fourth ribs and inserts into the anterior aspect of the medial scapular border.
 The lower component is the largest and most powerful, originating from the fifth through ninth ribs. It runs anterior to the scapula and inserts into the medial border of the scapula.
The lower component is the largest and most powerful, originating from the fifth through ninth ribs. It runs anterior to the scapula and inserts into the medial border of the scapula.
Levator Scapulae
Rhomboids
Humeral Propellers
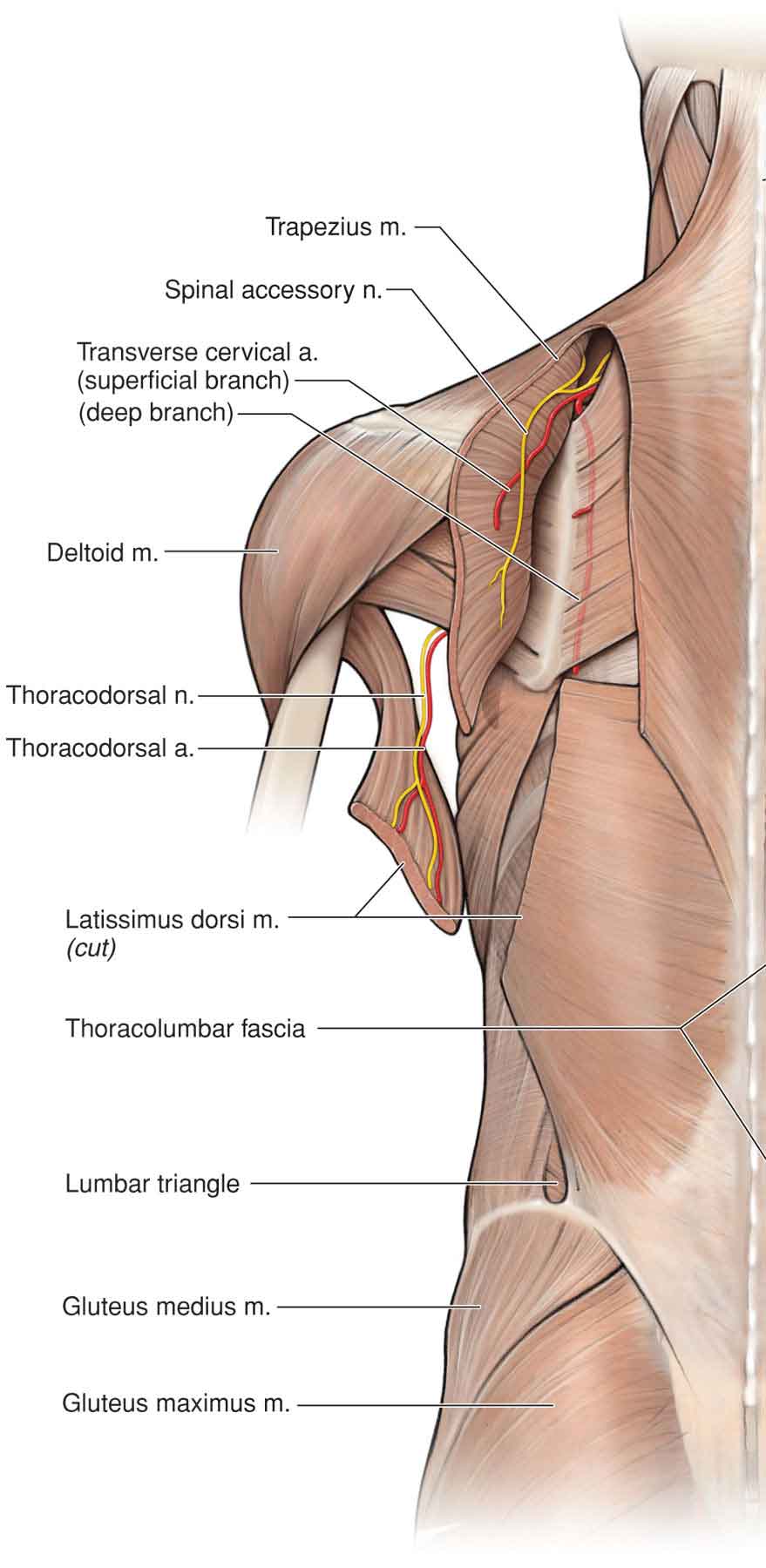
Latissimus Dorsi
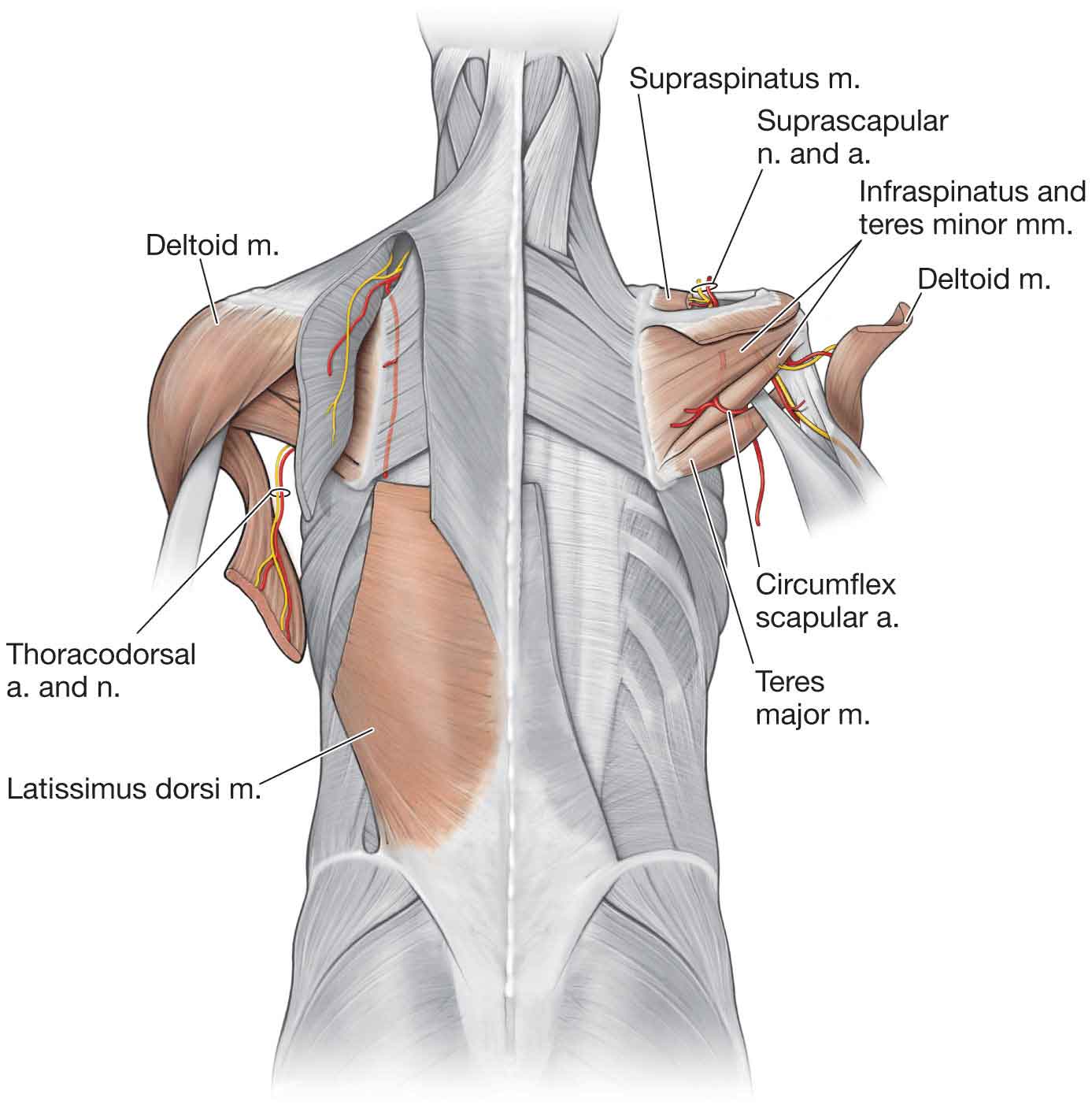
Teres Major
Pectoralis Major
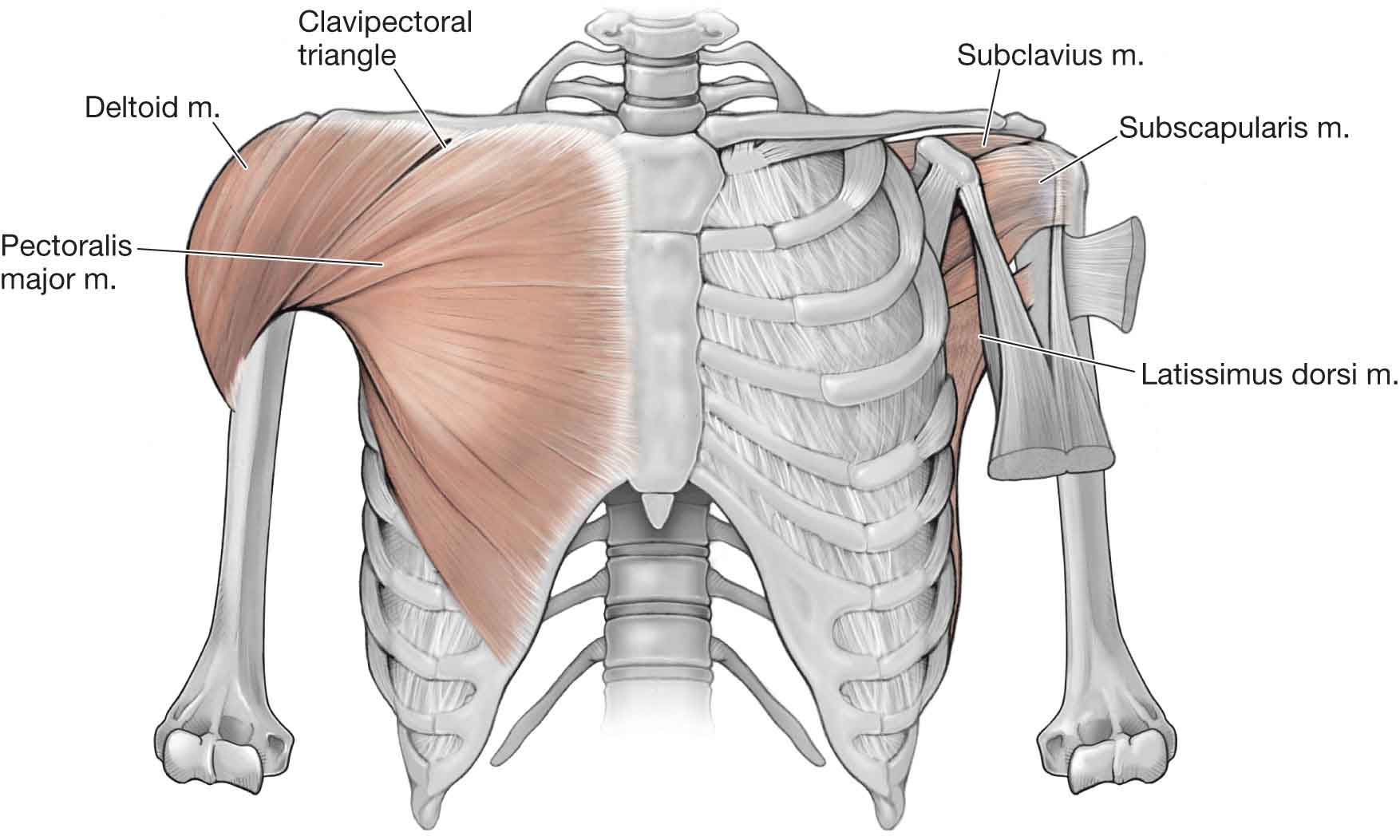
 Upper fibers (clavicular head)—IR, horizontal adduction, flexion, abduction (once the humerus is abducted 90 degrees, the upper fibers assist in further abduction), and adduction (with the humerus below 90 degrees of abduction) of the G-H joint.
Upper fibers (clavicular head)—IR, horizontal adduction, flexion, abduction (once the humerus is abducted 90 degrees, the upper fibers assist in further abduction), and adduction (with the humerus below 90 degrees of abduction) of the G-H joint.
 Lower fibers (sternal head)—IR, horizontal adduction, extension, and adduction of the G-H joint.
Lower fibers (sternal head)—IR, horizontal adduction, extension, and adduction of the G-H joint.
 The pectoralis major is innervated by the medial (lower fibers) and lateral (upper fibers) pectoral nerves (C8–T1 and C5–7, respectively).
The pectoralis major is innervated by the medial (lower fibers) and lateral (upper fibers) pectoral nerves (C8–T1 and C5–7, respectively).
Pectoralis Minor
Humeral Positioners
Deltoid
Shoulder Protectors
Rotator Cuff
 Assist in the rotation of the shoulder and arm. At the G-H joint, elevation through abduction of the arm (Table 16-3) requires that the greater tuberosity of the humerus pass under the coracoacromial arch. For this to occur, the humerus must externally rotate, and the acromion must elevate.117 ER of the humerus is produced actively by a contraction of the infraspinatus and teres minor, and by a twisting of the joint capsule. A force couple exists in the transverse plane between the subscapularis anteriorly and the infraspinatus and teres minor posteriorly in which cocontraction of the infraspinatus, teres minor, and subscapularis muscles both depresses and compresses the humeral head during overhead movements.
Assist in the rotation of the shoulder and arm. At the G-H joint, elevation through abduction of the arm (Table 16-3) requires that the greater tuberosity of the humerus pass under the coracoacromial arch. For this to occur, the humerus must externally rotate, and the acromion must elevate.117 ER of the humerus is produced actively by a contraction of the infraspinatus and teres minor, and by a twisting of the joint capsule. A force couple exists in the transverse plane between the subscapularis anteriorly and the infraspinatus and teres minor posteriorly in which cocontraction of the infraspinatus, teres minor, and subscapularis muscles both depresses and compresses the humeral head during overhead movements.Related posts:
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree





