Fig. 11.1
A schematic overview of the pathogenesis of intervertebral disc degeneration demonstrating the involvement of IL-1 in driving the aberrant cell biology and processes involved in matrix catabolism and generation of back pain
11.5.1 Soluble Regulators of Cellular Function
During degeneration, a range of pro-inflammatory cytokines and inflammatory mediators are increased. These include members of the interleukin family, including IL-1, IL-2, IL-6, IL-12 and IL-17, as well as interferon gamma (IFN-γ), TNF-α and the inflammatory mediators prostaglandin E2 (PGE2) and nitric oxide (NOx) (Kang et al. 1996; Olmarker and Larsson 1998; Le Maitre et al. 2005a, 2007b; Bachmeier et al. 2007; Akyol et al. 2010; Gabr et al. 2011; Studer et al. 2011). The pro-inflammatory cytokines are all thought to play independent roles in matrix catabolism, although interplays between the molecules have been identified. For example, IL-6 is thought to potentiate the response of nucleus pulposus cells to both IL-1 and TNF-α (Studer et al. 2011), while IL-17 synergises with both TNF-α and IFN-γ, increasing the catabolic activities of human nucleus pulposus and annulus fibrosus cells and possibly serving as a key regulator of inflammation in the degenerating disc (Gabr et al. 2011).
However, although there is evidence for the involvement of multiple pro-inflammatory cytokines in the pathogenesis of disc degeneration, the predominant catabolic cytokines appear to be interleukin-1 (IL-1) and tumour necrosis factor-alpha (TNF-α). While there is no clear consensus on which molecule mediates degeneration, research suggests both are fundamentally important in controlling the observed cellular and matrix changes.
11.5.1.1 Interleukin-1
Both isoforms of IL-1 (IL-1α and IL-1β) have been identified within the disc, along with their receptor (IL-1R1), the exported decoy receptor (IL-1RII) and their natural inhibitor (IL-1 receptor antagonist or IL-1Ra) (Le Maitre et al. 2005a). During degeneration, expression of IL-1α and β and IL1RI increases significantly in both the nucleus pulposus and inner annulus fibrosus. However, IL-1Ra expression does not increase and this imbalance leads to an excess of IL-1 isoforms in degenerate tissues. Importantly, in vitro studies have shown that IL-1 induces a number of cellular and molecular changes associated with disc degeneration. Stimulation of human nucleus pulposus cells with recombinant IL-1 has been shown to induce an upregulation of both MMPs, including MMPs 3 and 13, and ADAMTSs, including ADAMTS 4, a shift in collagen expression from II to I and reduction in aggrecan expression (Le Maitre et al. 2005a). There appear to be differences in the responses of normal and degenerate disc cells to IL-1 stimulation, with a more catabolic response in degenerate nucleus pulposus cells compared to normal. IL-1 stimulation also resulted in significant increases in both IL-1 isoforms by degenerate nucleus pulposus cells and a decrease in expression by normal nucleus pulposus cells, suggesting a homeostatic response in nondegenerate cells and an aberrant catabolic response once degeneration has been activated. IL-1 has also been shown to induce both angiogenesis (by inducing expression of VEGF) and neuronogenesis (via the stimulation of neurotrophic factors) into disc tissue (Lee et al. 2011) and stimulation of apoptosis (Cui et al. 2007; Zhao et al. 2007a). Conversely an inhibition or reversal of these processes has been demonstrated through the addition of exogenous IL-1Ra, and application of IL-1Ra has been proposed as a potential therapeutic intervention to inhibit intervertebral disc degeneration (Le Maitre et al. 2006a, 2007c; Box 11.1).
11.5.1.2 Box 11.1: Interleukin-1 as the Driving Force Behind the Pathogenesis of Disc Degeneration
While for some TNF-α has been the focus of investigation as the molecular regulator of disc degeneration, our research has focussed on the involvement of IL-1. These studies have demonstrated an increase in the expression of both isoforms (α and β) of IL-1, along with its receptor (IL-1RI) during degeneration. However, no such increase was demonstrated for its natural inhibitor, IL-1Ra, suggesting an imbalance that may be responsible for driving the cellular and matrix changes evident during degeneration. These roles include inducing expression of both MMPs and ADAMTSs, which are known to catabolise the extracellular matrix; reducing matrix component molecule expression, most notably aggrecan; inducing apoptosis and senescence of disc cells; and inducing both angiogenesis and innervation into the disc. However, the key question at present is: what initiates the upregulation of IL-1 that then induces the degenerative cascade? Elucidation of this mechanism may lead to the development of novel therapies or allow disc degeneration to be prevented.
11.5.1.3 TNF-α
TNF-α, like IL-1, has been shown to be capable of inducing neural ingrowths into the degenerate intervertebral disc. TNF-α has also been implicated in causing nerve root damage and sciatic pain, with blocking studies supporting this theory (Igarashi et al. 2000; Olmarker and Rydevik 2001). While evidence for a role for TNF-α in nerve ingrowth is compelling, evidence supporting its role in driving matrix catabolism during degeneration is less clear. Although expression of TNF-α is increased in degenerate tissues (Weiler et al. 2005; Bachmeier et al. 2007), other studies have either failed to identify TNF receptor I in degenerate samples or shown that there is no increase in the receptor gene expression in degenerate samples. The result suggests that native disc cells in vivo may not be able to respond to TNF-α (Le Maitre et al. 2007b). However, in studies where recombinant TNF-α was used to stimulate cultured nucleus pulposus cells, there were increases in expression of MMPs 1, 3, 9 and 13, as well as ADAMTSs 4 and 5, although induction of expression of a number of these enzymes was greater following stimulation with IL-1 than with TNF-α (Hoyland et al. 2008). Conversely, in situ zymography studies of normal and degenerate human nucleus pulposus tissue treated with either IL-1 or TNF-α indicated there was only an increase in enzyme activity in the IL-1 group and not the TNF-α group. In this study, addition of IL-1Ra caused a decrease in enzyme activity that was not evident following addition of anti-TNF. While demonstrating an increase in TNF-α in degeneration, the documented low expression of its receptor on disc cells together with differences in findings from the other studies begs the question: what is the target of nucleus pulposus cell secreted TNF-α? Does it play a more fundamental role in innervation and development of discogenic pain, than in matrix catabolism and tissue breakdown?
11.5.1.4 Anabolic Growth Factors
Growth factors have a number of effects on cells, most notably the shift in metabolic balance towards anabolism, promotion of cell proliferation and prevention of cell death. A range of growth factors, including many members of the TGF-β superfamily, are known to be present in the normal intervertebral disc, while evidence for their involvement in disease comes from both in vitro cell stimulation studies and studies of in vivo models of disc degeneration. Many workers have focussed on the ability of growth factors to stimulate proteoglycans synthesis by nucleus pulposus and annulus fibrosus cells and shown positive results following stimulation with transforming growth factor-β (TGF-β), epidermal growth factor (EGF), insulin-like growth factor-1 (IGF-1), connective tissue growth factor (CTGF) and the bone morphogenetic proteins (BMPs) 2, 7 (also known as osteogenic protein-1 or OP-1), 12, 13 (also known as growth and differentiation factor 6 (GDF-6) or cartilage-derived morphogenetic protein-2 (CDMP-2)) and 14 (also known as growth and differentiation factor 5 (GDF-5) or cartilage-derived morphogenetic protein-1 (CDMP-1)) either alone or in combination (Thompson et al. 1991; Osada et al. 1996; Gruber et al. 1997; Masuda et al. 2003; Tim et al. 2003; Imai et al. 2007a; Gilbertson et al. 2008; Le Maitre et al. 2009). Cell proliferation has also been demonstrated in vitro following stimulation with TGF-β and IGF-1, while the same growth factors and platelet-derived growth factor (PDGF) are all capable of reducing disc cell apoptosis and secretion of catabolic cytokines (Gruber et al. 1997, 2000).
In vivo studies also show positive effects of TGF-β, IGF-1, BMPs 2 and 5, fibroblast growth factor-2 (FGF-2) and OP-1 on cell proliferation, matrix synthesis and restoration of disc height in models of disc degeneration (Walsh et al. 2004; An et al. 2005; Masuda et al. 2006; Miyamoto et al. 2006). Of these, the most widely studied is OP-1, with positive effects noted both in vitro and in vivo. Indeed, OP-1 was capable of stimulating proteoglycans and collagen synthesis by human nucleus pulposus cells following IL-1 and chondroitinase ABC treatment in vitro (Takegami et al. 2005; Imai et al. 2007a) and restoring disc height and matrix degradation caused by annular needle injury or chemonucleolysis with chondroitinase ABC (Miyamoto et al. 2006; Imai et al. 2007b). OP-1 also reduced the production of aggrecanase, MMP-13, substance P, TNF-α and IL-1β, suggesting that it may have both anabolic and anti-catabolic effects (Chubinskaya et al. 2007). The effect of OP-1 on substance P expression, combined with the interplay between catabolic cytokines and pain markers or pain modulators, has led to the hypothesis that application of recombinant growth factors may have beneficial effects in terms of pain reduction, as well as matrix restoration during degeneration (Kawakami et al. 2005). However, the clinical translation of growth factor therapies is complicated by their diverse and sometimes biphasic roles. While growth factors such as TGF-β and CTGF are known to promote extracellular matrix synthesis, studies on clinical human samples have linked their over-expression to fibrosis and angiogenesis (Ali et al. 2008; Peng et al. 2009). The application of growth factor therapies is further complicated by possible changes in receptor distribution. While data in this area is limited, studies suggest that there is no change in the expression levels of major growth factor receptors, including TGFβRII, BMPRII, FGFR3 and IGFRI (Le Maitre et al. 2005b; Peng et al. 2006). However, expression of growth factor receptors, including TGFRII, FGFR3, IGFRI and VEGF receptors I and II, on ingrowing blood vessels (Haro et al. 2002; Le Maitre et al. 2005b) and in granulation tissue in painful degenerate discs (Peng et al. 2006) suggests that the use of growth factors should carefully be controlled to avoid stimulation of unwanted events such as angiogenesis. Therefore, a more detailed understanding of their expression profiles and roles in both disc degeneration and repair are currently required.
Another area where growth factors show potential is in the stimulation of adult mesenchymal stem cell (MSC) differentiation. Previous studies have shown that TGF-β, along with members of the BMP family, most notably BMPs 2 and 14, can stimulate MSC differentiation towards nucleus pulposus-like cells in vitro (Stoyanov et al. 2011; McCanless et al. 2011). Given the increasing information on the nucleus pulposus phenotype, further studies will no doubt shed light on the role that growth factors play in MSC differentiation and may lead to combined regenerative cell/growth factor therapies for treatment of disc degeneration.
11.6 Changes in Disc Nutrition and Oxygen Tension
As mentioned previously, nutrient supply to the disc is predominantly from blood vessels in the vertebral bodies and occurs via diffusion through the cartilaginous end plates. This theory has been well studied and confirmed using a range of tracer diffusion experiments, including MRI contrast media, fluorescent and radioactive tracers and gaseous tracers such as nitrous oxide (Brodin 1955; Holm and Nachemson 1982, 1983; Adams and Hutton 1986; Urban et al. 2001). Exposure to cigarette smoke has also been shown to inhibit transport of oxygen into, and lactic acid out of, the disc in animal experiments due to constriction of the microvasculature in the vertebrae (Holm and Nachemson 1988); this finding supports epidemiological evidence linking smoking to disc degeneration in humans. The size and charge of solutes also affect their penetration into the disc, with anions showing lower rates of diffusion than cations due to the polyanionic nature of the disc (Urban et al. 2004), while larger molecules such as albumin are effectively prevented from diffusing through the cartilaginous end plate (Urban et al. 2004). Compared to uni- or bivalent electrolytes, and the fact that glucose is a relatively large molecule, Urban et al. opined that diffusion into and through the disc may be slow (Urban et al. 2004). This lack of nutrient supply to the core of the nucleus pulposus is reflected in the non-uniform cell distribution throughout the disc: cell number in the outer annulus (which is closer to capillaries surrounding the tissue) is substantially higher than that of the nucleus which can be as far as 8 mm from the nearest capillary.
With age, the nutrient supply to the disc is reduced in part due to reductions in both the density and integrity of capillaries in the vertebral bodies and in part due to calcification of the cartilaginous end plates (Bernick and Cailliet 1982; Roberts et al. 1996). While it is currently unclear whether cartilaginous end-plate calcification is causative of, or the result of, disc degeneration, it is thought to play an important role in disease progression by posing a significant barrier to diffusion of solutes into and out of the disc. However, recent μCT studies on graded normal and degenerate human samples have suggested that, contrary to popular belief, porosity in vertebral end plates increases in degeneration by as much as 130 %, while trabecular thickness decreases by as much as 50 % (Rodriguez et al. 2011, 2012). One result of these changes appeared to be an increase in cell proliferation and decrease in proteoglycan content in the nucleus pulposus. While solute transport was not assessed in this study, the authors proposed that ischemic cell changes in degeneration may reflect capillary transport activity rather than a decrease in end-plate permeability.
Whatever its cause, a reduction in essential nutrients is thought to drive the progression of degeneration. Given that the main energy-generating pathway in disc cells, even in the presence of oxygen, is glycolysis (Holm et al. 1981; Ishihara and Urban 1999), which requires principally glucose and produces lactic acid, free diffusion of solutes to and from the cells is essential. Evidence suggests that reduction in glucose concentration below 0.5 mmol/L, even for a relatively short period, can cause cell death. Likewise, reduction in the pH to below 6.4 can also promote death (Horner and Urban 2001; Bibby and Urban 2004), while less severe reductions in pH can impact cell metabolism (Ohshima and Urban 1992). Evidence from a range of assays, including biochemical assays and microelectrode measurements, shows that this drop in glucose concentration and pH is similar to that observed in degenerate discs. Therefore, while cell activity and even viability may be impaired by a decrease in nutrient supply, enzyme activity is not reduced. In this case, there would be an imbalance between matrix anabolism and catabolism which may contribute to an elevation in matrix degradation in degenerate discs.
The other key metabolite in the disc is oxygen, although its role is less clear than that of glucose. Oxygen levels vary widely in human discs, with no clear correlation between a change in oxygen tension and disc degeneration. However, there exists a steep oxygen gradient within discs – studies in dogs demonstrating a decrease from 8 to 10 % O2 at the disc-vertebral body interface to 0.3–0.5 % in the centre of the nucleus pulposus (Holm et al. 1981). Human discs show similar trends, with readings as low as 0.7 % O2 in the core of the degenerate human nucleus (Bartels et al. 1998). While oxygen is consumed by disc cells, relatively little CO2 is produced and disc cells can survive for at least 2 weeks without oxygen (Horner and Urban 2001), suggesting oxidative phosphorylation is not the primary mechanism for energy metabolism. Further studies on both canine and bovine disc cells demonstrated that as O2 concentration decreases from 21 to 1 %, there was a decrease in oxygen consumption of around 75 % and a sharp increase in lactate production (Ishihara and Urban 1999). This suggests a positive Pasteur effect since glycolysis is stimulated under hypoxic conditions. However, studies on the effect of hypoxia on disc cells have also demonstrated cellular inactivity (Horner and Urban 2001) and a loss of matrix synthesis (Ishihara and Urban 1999) below an oxygen concentration of around 5 %. As the increase in lactate production under hypoxic conditions is likely to reduce the pH, this in turn would reduce matrix synthesis (Ohshima and Urban 1992) and eventually cause cell death (Horner and Urban 2001; Bibby and Urban 2004). Hence, the effects of low oxygen and low pH are likely to be cumulative and a destructive influence of disc cell survival. The mechanism by which cells sense oxygen and accommodate to the oxemic state is discussed in exhaustive detail in Chap. 6.
Confounding the problem of evaluating the consequences of oxygen and nutrient limitation is the fact that human tissues are different from those of animal models. Therefore, investigators have increasingly turned to mathematical finite element modelling to elucidate the effect of nutrient limitation on the disc microenvironment and cellular metabolism. However, since all of the variables involved in disc nutrition are poorly understood, the studies published in this area are relatively simple, dealing with only one or two aspects of nutrient supply. Nevertheless, they all suggest that limited nutrient supply affects disc cell viability and metabolic activity (Selard et al. 2003; Yao and Gu 2006; Mokhbi et al. 2009; Malandrino et al. 2011; Jackson et al. 2011).
As with findings from finite element modelling, in vitro studies appear to confirm that during degeneration, glucose limitation and decrease in pH are the predominant factors affecting cell metabolism. While nucleus pulposus cells have mechanisms to regulate intracellular pH, such as the expression of the carbonic anhydrases 9 and 12 (Minogue et al. 2010a), our preliminary data indicates that the expression of these molecules decreases with degeneration, suggesting that cells may not be capable of withstanding the low pH over extended periods (Fig. 11.2). Likewise, changes in the nucleus pulposus membrane glucose transporters (GLUTs), present in degenerative tissues, suggest that there are molecular adaptations to compensate for the reduced glucose concentrations (Richardson et al. 2008b). However, the harsh environment of the degenerate disc is likely to be detrimental not just to resident cells but also to cells that are introduced into the disc. Accordingly, for future cell-based therapies, the introduction of too many cells, or cells which are unable to withstand the conditions of the microenvironmental niche, may exacerbate the degenerative problem rather than provide a cure.
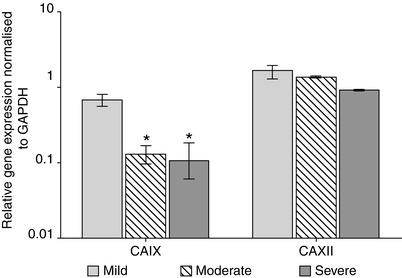
Fig. 11.2
Histogram illustrating real-time quantitative PCR data demonstrating a decrease in expression of carbonic anhydrase 9 (CAIX) and 12 (CAXII) in nucleus pulposus cells during disc degeneration. The expression of CAIX is significantly decreased with disease progression, while a decrease is also seen for CAXII. The carbonic anhydrases play a role in regulating intracellular pH, and a downregulation in their expression may result in cells being incapable of withstanding the harsh physicochemical environment of the degenerate intervertebral disc
11.7 Cell Ageing and Death
During development, the nucleus pulposus is populated by large, vacuolated, metabolically active and morphologically distinct notochordal cells which produce high levels of proteoglycans (Boos et al. 2002; Cappello et al. 2006). However, by the age of 10, this population has been replaced by smaller, chondrocyte-like nucleus pulposus cells which are less metabolically active (Wolfe et al. 1965; Pazzaglia et al. 1989; Boos et al. 2002; Guehring et al. 2008). The period between 3 and 10 years of age, during which identifiable notochordal cells disappear, is also the time during which there is a high level of cell death (Boos et al. 2002). These changes appear to signal the initiation of a transition from a highly hydrated, gelatinous extracellular matrix to a more fibrous, cartilaginous nucleus pulposus seen in adults. It also coincides with the earliest identifiable signs of degeneration seen in MRI studies. In addition, histological studies have identified matrix changes in this age group, which progress throughout adult life (Boos et al. 2002). Although these changes may be considered to be part of the normal ageing process, as discussed previously, the accelerated matrix degradation suggests that in disc degeneration, there is premature ageing.
Several studies have identified necrotic cells within the disc which increase both with age and degeneration; however, more recently, apoptosis has been identified as the principle mechanism of cell death. Evidence for this has come from a number of studies, using a diverse range of markers including transferase-mediated dUTP nick-end labelling (TUNEL) staining (Gruber and Hanley 1998; Lotz and Chin 2000; Rannou et al. 2004; Kim et al. 2005; Risbud et al. 2005; Heyde et al. 2006; Park et al. 2006; Loreto et al. 2011), annexinV-propidium iodide flow cytometry (Rannou et al. 2004; Risbud et al. 2005; Park et al. 2006), caspase activity analysis (Rannou et al. 2004; Heyde et al. 2006; Park et al. 2006; Tschoeke et al. 2008) and gene expression studies of apoptosis-related markers such as Bax and Bcl-2 (Heyde et al. 2006; Tschoeke et al. 2008; Loreto et al. 2011). The method employed to establish the incidence of apoptosis appears to influence findings both in vivo and in vitro. Serum deprivation resulted in only 1 % cell apoptosis when studied using TUNEL staining (Gruber et al. 2000); in contrast, similar conditions increased in the incidence of apoptosis in rat annulus fibrosus cells to 56 % when annexinV-propidium iodide flow cytometry was used (Risbud et al. 2005). Other studies have also suggested that incidence of apoptosis in disc cells can be as high as 73–74 % (Gruber and Hanley 1998; Ha et al. 2006), although this would be an overestimate since it would result in the de-cellularisation of the disc within days (Alvarez and Ortiz 1999). Conversely, studies using fluorescent cell viability assays on fresh disc tissue have repeatedly demonstrated 60 % to over 90 % cell viability even in degenerate and scoliotic discs, further suggesting that the apoptosis measurements are an overestimation (Bibby et al. 2002; Johnson and Roberts 2007). However, while no causative link has been found between apoptosis and increased extracellular matrix degradation, it is probable that programmed cell death plays an essential role in the pathogenesis of disc disease.
Autophagy, a pathway that may lead to programmed cell death, has recently been identified in the rat nucleus pulposus (Ye et al. 2011) and annulus fibrosus (Shen et al. 2011). We have also demonstrated evidence of autophagy in tissues of the degenerate human intervertebral disc, through immunostaining for the key markers LC-3 and beclin-1 (Fig. 11.3). While autophagy can lead to cell death, it can also maintain cell viability during periods of environmental or nutritional stress by catabolism of intracellular organelles and unfolded or damaged proteins. Therefore, more work is required to establish the role of autophagy in disc degeneration.
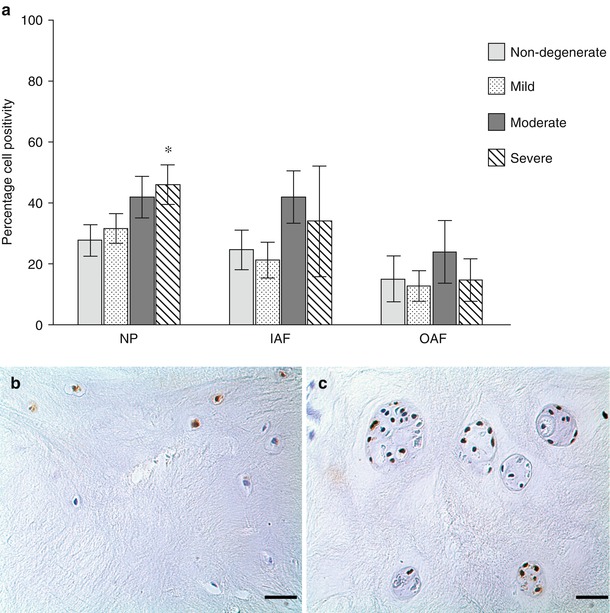
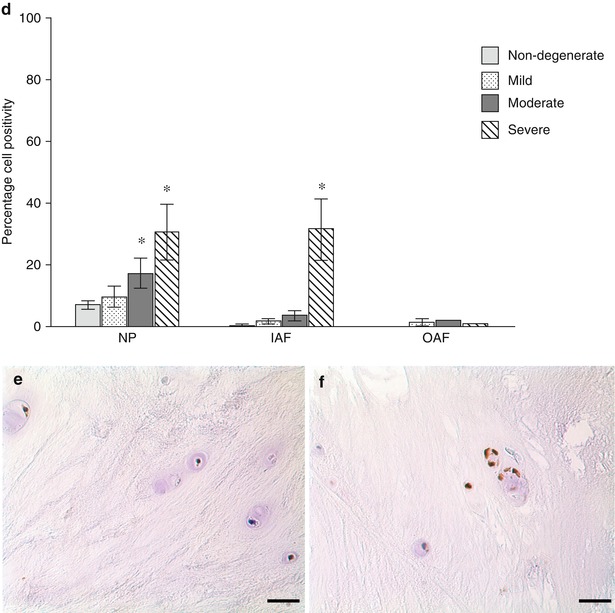
Fig. 11.3
Evidence for the presence of autophagy markers in normal and degenerate human IVD cells. Immunohistochemistry for LC-3 (a–c) and beclin-1 (d–f) demonstrated staining for both markers in normal (b, e) and degenerate (c, f) nucleus pulposus cells, with strong immunopositivity in degenerate cell clusters. Semi-quantitative analysis of cell positivity demonstrated significant increases in both LC-3 and beclin-1 in the nucleus pulposus with progression of degeneration (a, d) and in the inner annulus fibrosus (IAF), but not outer annulus fibrosus (OAF), for beclin-1. Although preliminary data, these findings suggest a role for autophagy in disc cells and that there may be an increase in the number of autophagic cells in intervertebral disc degeneration
Studies on intervertebral disc cell number suggest that rather than a steady decline in cell numbers over time, there is a cyclical pattern of cell death followed by a period of proliferation. Indeed between 11 and 16 years, following the extensive cell death noted between 3 and 10 years, there is notable cell proliferation. The authors hypothesise that this is a compensatory mechanism in response to the early matrix changes seen during this time period (Boos et al. 2002). Cell clustering and proliferation, a characteristic phenomenon of degenerating tissue, is seen in the nucleus pulposus; both parameters increase both with age and with stage of degeneration (Boos et al. 2002). Both histological parameters are routinely identified adjacent to clefts and tears within the tissue, possibly due to a localised increase in the local nutrient supply in these regions (compared to an overall decrease in nutrients within the disc) rather than as a reparative mechanism (Beard et al. 1981; Boos et al. 2002; Zhao et al. 2007b). This hypothesis is further supported by evidence obtained from scoliotic discs which exhibit an increase in cell population in the inner annulus fibrosus in association with regional neovascularisation (Beard et al. 1981). Although signs of cell death are seldom seen, markers of cell proliferation, most notably proliferating cell nuclear antigen (PCNA) and the proliferation-associated Ki-67, have been identified within these cell clusters (Johnson et al. 2001; Zhao et al. 2007b).
A number of potential theories have been proposed to explain this finding. The increased local nutrition may enable cells within clusters to resist apoptosis, or possibly dead cells may be cleared more quickly from clusters than from the rest of the disc. In this respect, other workers have identified morphologically nucleus pulposus-like but CD68-positive cells within cells cluster in the nucleus pulposus of degenerate discs (Nerlich et al. 2002). These cells have the ability to phagocytose apoptotic bodies and are thought to be transformed nucleus pulposus cells, rather than infiltrating cells. In vitro studies have also demonstrated the ability of bovine nucleus pulposus cells to phagocytose apoptotic cells, suggesting that cell clearance from the disc may be undertaken by resident disc cells rather than infiltrating macrophages or monocytes (Jones et al. 2008).
Within clusters, increased proliferation predisposes the resident cells to replicative senescence. Several studies have identified markers of cellular senescence in degenerate discs (Roberts et al. 2006a; Gruber et al. 2007), and in 2007, a link between accelerated cell senescence and degeneration was identified. Through both gene and protein expression studies, Le Maitre et al. (2007a) showed that there was an increase in expression of p16INK4A, a cell cycle inhibitor which is upregulated in senescence, a decrease in both mean telomere length and replicative potential and an increase in senescence-associated β-gal staining. While similar trends were identified with ageing, these senescence changes were positively related to stage of degeneration, irrespective of age, clearly demonstrating a role for senescence in degeneration. Such findings are supported by those of Kim and colleagues who also demonstrated an accumulation of senescent cells in degenerate discs. These workers reported that the telomere-based p53-p21-pRB pathway plays an important role in inducing senescence in nucleus pulposus cells (Kim et al. 2009). In addition to replicative senescence, caveolin-1, a marker of stress-induced premature senescence (SIPS), has also been identified in the disc. Its expression is correlated with increases in p16INK4A, but not with age, suggesting a potential role for SIPS in degeneration (Heathfield et al. 2008). Interestingly, various stressors such as reactive oxygen species (Homma et al. 1994; Chen et al. 1995), mechanical loading (Martin et al. 2004) and the presence of cytokines such as IL-1 (Dai et al. 2006), all thought to play a role in the processes leading to disc degeneration, have been reported to induce SIPS, suggesting that this type of senescence may contribute to disc degeneration. However, more work is required to confirm these links.
Expression of cell senescence markers in the study by Le Maitre et al. was also shown to be correlated with changes in expression of two proteolytic enzymes, MMP 13 and ADAMTS 5, which are known to be upregulated in degenerate tissue. This finding indicated that a link may exist between senescence and induction of matrix catabolism (Le Maitre et al. 2007a). As well as inhibiting proliferation, studies in other tissues show that senescent cells adopt an altered phenotype, described as the senescence-associated secretory phenotype (SASP) (Freund et al. 2011). Such cells secrete a range of pro-ageing and catabolic factors, most notably IL-1, which are found to be elevated in the degenerate intervertebral disc. Studies on senescent chondrocytes show similar increases in cytokines, as well as MMPs (including MMP 13) and other proteolytic enzymes. Likewise, these cells exhibit a decreased response to anabolic stimuli that is characteristic of cells from degenerate discs. While more work is clearly needed to elucidate a potential causative link between senescence and disc degeneration, it is clear that the increase in senescence in degeneration and the phenotype adopted by the senescent cell suggest that this process may be important in disease progression.
11.7.1 Box 11.2: Elucidation of Cellular Phenotype Is Central to Improving Understanding of the Degenerative Process and Development of Novel Therapies
The adult nucleus pulposus is populated by cells routinely described as ‘chondrocyte-like’ based on their rounded morphology and expression of SOX-9, type II collagen and aggrecan, although controversy has long surrounded their origin and exact phenotype. However, recent microarray studies from our group and others have begun to elucidate the true phenotype of these cells. They reveal interesting gene signatures, the significance of which are yet to be fully understood. However, the expression of some of these genes (such as FOXF1) lends weight to the growing body of evidence from developmental biology studies and other sources that the adult human nucleus pulposus is populated, at least in part, by notochordally derived cells. Notochordal cells produce higher levels of proteoglycans than mature disc cells and animals which retain their notochordal cells have a gelatinous nucleus pulposus which does not show signs of degeneration. How the novel marker genes change during degeneration and their potential role in the pathogenesis of disease has yet to be fully elucidated. However, the elucidation of the nucleus pulposus phenotype has important implications for the development of novel stem cell-based regenerative medicine therapies as it allows researchers to understand the end point of differentiation and avoid generation of chondrocyte-like cells which may not produce a correctly functioning extracellular matrix.
11.8 Response to Mechanical Load
The human intervertebral disc is exposed to a number of physical stresses, including compressive loading, which predominantly affects the nucleus pulposus, and stretch, shear and torsion which mainly affect the annulus fibrosus. Using pressure-sensitive needles inserted into the nucleus pulposus, Nachemson et al. demonstrated that loads experienced within the human disc ranged from 250N when lying down to 1900N when lifting a 10 kg weight with a bent spine (Nachemson 1981). However, further analysis of this data, taking into consideration the tensile forces exerted by muscles in the back, suggests that loading may be as high as 9000N when lifting. It has been suggested that this combination of compression and flexion is responsible for disc prolapse, with over-flexion combined with moderate load being more detrimental than excessive load with moderate flexion (Hutton and Adams 1982). Following on from Nachemsons research, Wilke et al. demonstrated a load of between 0.1 MPa when lying prone and 2.3 MPa when lifting a 20 kg weight with a flexed spine (Wilke et al. 1999); however, this study again did not take into consideration the tensile forces exerted by the musculature in the back.
The investigations mentioned above indicate that cells in the disc experience substantial loads, and predictably, these loads are thought to have a profound effect on cell behaviour. Indeed, mechanical-loading studies using in vivo animal models and in vitro cell culture techniques have demonstrated that the type, magnitude, frequency and duration of loading are paramount in determining cell response (MacLean et al. 2004, 2005; Wang et al. 2007; Wuertz et al. 2009; Korecki et al. 2009; Sowa et al. 2011). In particular, while moderate loads and low-frequency loading both promote anabolic responses, high-magnitude, high-frequency as well as sustained static loads all elicit a catabolic or anti-anabolic response in disc cells. Using human intervertebral disc cells, Neidlinger-Wilke et al. reported increased matrix protein expression (type I collagen and aggrecan), but no change in the expression of matrix-degrading enzymes (MMPs 1, 2, 3 and 13) after low-magnitude compressive loading; high-magnitude loads led to decreased matrix protein expression with increased matrix-degrading enzyme expression (mainly MMP 3) (Neidlinger-Wilke et al. 2006). Similarly, Handa et al. found that load influenced proteoglycan synthesis and MMP gene expression in human nucleus explants, with low loads promoting matrix anabolism and high loads leading to matrix catabolism (Handa et al. 1997).
While the effect of mechanical load on nondegenerate disc cells has been well documented, studies into the response of degenerate nucleus pulposus cells remain limited. To address this need, recent investigations have compared loading responses of nondegenerate and degenerate human nucleus pulposus. With physiological loads, Le Maitre et al. showed that nondegenerate nucleus pulposus cells produced an anabolic response, while degenerate cells remained unresponsive (Le Maitre et al. 2008). This finding suggests an alteration in mechanotransduction pathways between normal and degenerate cells. In support of this notion, Gilbert et al. showed differences in response to cyclic tensile strain between nondegenerate and degenerate human annulus fibrosus cells (Gilbert et al. 2010, 2011). Accordingly, while cyclic tensile strain applied at 1 Hz to nondegenerate annulus fibrosus cells resulted in a decrease in catabolic gene expression, the same strain caused a decrease in anabolic gene expression by degenerate annulus fibrosus cells. In the latter cells, there was evidence of an altered mechanotransduction pathway which appeared to be independent of cytokine involvement (Gilbert et al. 2010, 2011). While the implications of these changes require further elucidation, the profound effects of mechanical forces on cell behaviour cannot be ignored, and the possibility exists that force plays a fundamental role in the initiation or progression of disc degeneration. For a further discussion of the effects of force on cells, see Chap.7.
11.9 Clinical Implications: Relevance of Understanding the Cell Biology and Pathogenesis of Intervertebral Disc Degeneration for Development of Novel Therapeutic Agents
Current clinical interventions for back pain are predominantly aimed at relieving symptoms rather than treating the underlying disorder. In many cases, this involves regular administration of pain-relieving pharmaceuticals, e.g. nonsteroidal anti-inflammatories, or application of more novel therapies, such as transcutaneous electrical nerve stimulation, physical manipulation, exercise therapy or behavioural therapies (Bogduk 2004). However, despite widespread use, the efficacy of these interventions is still questionable (van der Roer et al. 2005). In patients who are unresponsive to conservative therapies, but have identifiable imaging deficits and clinical symptoms of back pain, fusion surgery is the ultimate end point (Errico 2005). This approach removes the source of pain, but due to alterations in spinal biomechanics, it reduces mobility and can cause problems at adjacent motion segments (Hilibrand and Robbins 2004). One alternative to spinal fusion is whole intervertebral disc or nucleus pulposus transplantation, using either autologous or allogeneic tissues (Katsuura and Hukuda 1994; Luk et al. 1997). These procedures have been successfully performed, but require complicated surgery, and issues have arisen regarding loss of tissue integrity, tissue instability and immunogenicity (Alini et al. 2002). While tissue transplantation does not seem a feasible alternative, an increasing range of disc replacement devices are currently being investigated. These include devices such as the prosthetic disc nucleus (PDN) device or whole-disc replacements such as Charite and ProDisc (Jin et al. 2003; Guyer et al. 2009; Delamarter et al. 2011). Large-scale trials are ongoing with these devices, and with the whole-disc replacement, a significant reduction in pain score has been demonstrated. However, complications including device migration, extrusion and failure are all issues and studies have so far failed to show improved outcomes compared to fusion (Errico 2005; Di et al. 2005; Lindley et al. 2010). For a more detailed discussion of these devices, see Chaps. 13 and 14.
This lack of clinically successful long-term treatment for discogenic back pain has led researchers to investigate both biological modulators of disc cell function and novel cell-based tissue engineering and regenerative medicine therapies.
In line with the increase in knowledge surrounding the control of disc matrix anabolism and tissue degradation during degeneration, the utility of a number of biologically active agents has been evaluated. These modulators include cytokine inhibitors, such as IL-1Ra, to inhibit matrix degradation (Le Maitre et al. 2006a) and growth factors, such as OP-1, to promote matrix restoration (Masuda et al. 2006). Given the growing understanding of nerve ingrowth into the degenerate disc, it should also be possible to use biological modulators to prevent or inhibit migration of nerves into the disc and thereby block transmission of discogenic pain. Of course, while biological agents provide a mechanism to inhibit the progression of early-stage degeneration, they may not be sufficient to regenerate tissue at later stages of degeneration. At this late stage, pain is the driver behind an individual seeking clinical help, and thus, identification of a suitable cohort of early-stage patients may be difficult. However, a clearer understanding of genetic predisposition may enable individuals to be screened and interventions targeted prior to the development of symptomatic back pain.
The other area where biological modulators may be beneficial is in combination with cell-based tissue engineering therapies, where they may be used to stimulate cell differentiation or matrix formation. For intervertebral disc tissue engineering, the use of nucleus pulposus cells would initially appear to be the obvious choice, with disc cell reimplantation showing promising results in both animal models and small-scale human safety trials (Meisel et al. 2006). However, the alterations in disc cell phenotype and function during degeneration raises questions about the applicability of using autologous cells, isolated from degenerate discs, for such therapies. Problems also surround the acquisition of autologous cells from discs adjacent to the degenerate level, as the local damage caused by removing tissue from those regions has been shown to lead to degeneration at an accelerated rate (Nomura et al. 2001). The use of allogeneic cells from young, healthy donors also poses immunogenicity risks. For these reasons, the focus has shifted to the use of autologous adult stem cells, derived from either bone marrow or adipose tissue, which have been shown to be capable of differentiating into nucleus pulposus-like cells and producing an nucleus pulposus-like extracellular matrix both in vitro and in vivo (Sakai et al. 2005; Richardson et al. 2006a, b, 2008a; Box 11.2). The potential value of these therapies is that they may be able to regenerate disc tissue and restore long-term functionality. However, for the therapy to be successful, it is important to take into account the environment into which these cells will be implanted. Thus, these studies must be conducted under conditions which closely mimic the complex microenvironment of the degenerate disc. This microenvironment includes increased levels of catabolic cytokines, with low levels of nutrients and a low pH, all factors which are known to affect cell function and may have a profound effect on the ability of stem cells to survive, differentiate or secrete matrix.
To aid cell survival and function following implantation, advanced biomaterials are required which can be implanted using minimally invasive procedures. These materials must be deformable and able to withstand the loads experienced by the spine; they must be able to support or promote cell survival, differentiation and matrix formation; and finally, they can biodegrade over a suitable timescale to non-toxic by-products. At present, no ‘ideal’ biomaterials exist, but the field is developing rapidly and cell-based regenerative medicine therapies appear likely to revolutionise the treatment of discogenic back pain over the coming decades.
11.10 Summary of Critical Concepts Discussed in the Chapter
Disc degeneration is a complex, multifactorial process, in which disc cells themselves play a fundamental role. While there are strong genetic predeterminants, a clear predisposition is difficult to detect.
During degeneration, there is a cell-driven loss of proteoglycans from the extracellular matrix, which results in gross morphological, biological and biomechanical changes within the spine and the development of clinical back pain.
Members of the MMP and ADAMTS families are responsible for breakdown of the extracellular matrix, including an imbalance between ADAMTSs and TIMP that could lead to aggrecan degradation.
In the disease state, the loss of aggrecan, with the shift to versican production, reduces the disc water content and the shift to collagen type I production results in a more fibrous tissue, less capable of withstanding load.
Expression levels of NGF and BDNF are increased in individuals with symptomatic disc degeneration. The disc cells express the high-affinity NGF and BDNF receptors and the low-affinity NGF/BDNF receptor p75NTR, as well as SP, suggesting autocrine signalling.
Interplay between catabolic cytokines and neurotrophins, neurotrophin receptors and chemorepellant molecules may guide nerve ingrowth during degeneration.
The predominant catabolic cytokines are IL-1 and TNF-α. During degeneration, interleukin family members, interferon gamma (IFN-γ), TNF-α, PGE2 and NOx are increased.
TGF-β, along with BMPs 2 and 14, can stimulate MSC differentiation towards nucleus pulposus-like cells in vitro.
A reduction in essential nutrients drives degeneration. With tissue hypoxia, an increase in lactate production reduces the pH, which in turn reduces matrix synthesis and can cause cell death. The effects of low oxygen and low pH may be cumulative and promote disc cell death.
During 3 and 10 years of age, identifiable notochordal cells disappear, and the highly hydrated, gelatinous extracellular matrix converts to a more fibrous, cartilaginous nucleus pulposus.
It is probable that apoptosis plays a critical role in the pathogenesis of disc disease. Autophagic changes have also been identified in the degenerate disc. While autophagy can lead to cell death, it can also maintain cell viability during periods of stress.Related posts:
 The Role of the ADAMTS Proteins in the Intervertebral Disc
Development of the Intervertebral Disc
Introduction to the Structure, Function, and Comparative Anatomy of the Vertebrae and the Intervertebral Disc
Tissue Engineering of the Intervertebral Disc
Use of Stem Cells for Regeneration of the Intervertebral Disc
Epidemiology of Lumbar Disc Degeneration
The Role of the ADAMTS Proteins in the Intervertebral Disc
Development of the Intervertebral Disc
Introduction to the Structure, Function, and Comparative Anatomy of the Vertebrae and the Intervertebral Disc
Tissue Engineering of the Intervertebral Disc
Use of Stem Cells for Regeneration of the Intervertebral Disc
Epidemiology of Lumbar Disc Degeneration

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


