CHAPTER 46 Developmental and Functional Anatomy of the Cervical Spine
INTRODUCTION
A thorough knowledge of human anatomy continues to be the cornerstone for all diagnosis and treatment planning. This awareness allows spine physicians to rise above mediocrity and arm themselves for the complex issues involved in spine care. Unlike peripheral joints, spinal anatomy involves multiple joints interacting in conjunction to bring about complex, multiplanar motions. This motion has physiologic restrictions and weaknesses based upon anatomy. These weaknesses allow for the unfortunate consequences resulting in axial and radicular pain syndromes that spine physicians know so well. This chapter will provide the reader with a basic understanding of both the developmental and functional anatomy of the cervical spine.1
DEVELOPMENTAL ANATOMY
Embryologic development
In order to give a temporal flow to the embryologic development of the cervical spine we will use the general staging scale devised originally by Streeter.2 This scale consists of 23 stages of growth that starts with fertilization and runs to the 60th day of growth.3 Each stage typically has a duration of 2–3 days in length.4 Growth and differentiation of the neurospinal axis begins in embryonic stage 6. These cells typically originate from the primitive streak and node at the 13th–15th day from fertilization. In the following stage the cells that will form the notochord travel rostrally from the primitive node. The notochordal cells’ growth causes abutment to the overlying ectoderm which thickens and becomes the neural plate.5,6 This neural plate serves as the primordium for all nervous system evolution.7 This stage occurs at the 17th–19th day (stage 8) and marks the first stage of nervous system development. The superior notochord promulgates growth of neuroectodermal structures, while the inferior notochord stimulates growth of mesodermal structures.8 In stage 9 the neural plate molds itself into a neural fold/groove.6 This stage is also notable as the first three somites are formed during this period of the 19th to 21st day. The somites represent paired, blocklike masses of mesoderm lying alongside the neural tube. The human embryo develops 42–44 paired somites; 4 occipital, 8 cervical, 12 thoracic, 5 lumbar, 5 sacral, and 8–10 coccygeal somites. They consist of two cell collections: the ventromedially placed sclerotomal cells and dorsolaterally situated dermomyotomes.9 The sclerotomal structures are responsible for the formation of the vertebral column, and the dermomyotomal cells form the segmental skin and musculature.10 It is postulated that early vertebral segmentation occurs during this stage; most notable is the formation of the first two cervical vertebrae and cranial–vertebral junction.6 These bony structures are formed by the first eight occipital somites. Spinal cord primordium is also present during this stage with continuing growth occurring through stage 12. In this stage the inferior neuropore closes and 21 to 29 somite pairs have formed. At this point, vertebral differentiation has occurred through the lumbar segments. Stage 13 (days 28–32) marks completion of pontine and cervical flexures of the spine and continuing neurulation caudal to closure of the posterior neuropore.11 Contouring of the caudal neural tube and adjacent vertebrae begins in this stage and progresses through gestation and into early infancy.6,11 Structures such as the conus medullaris, ventricular terminalis, and filum terminale originate from this caudal neural tube.12
Specific vertebral development occurs through three stages: membrane, cartilage, and bone formation.13 Membrane formation occurs as the notochord encounters the neural tube dorsally and the foregut ventrally. As growth progresses, cavities are formed between the notochord and neural tube/foregut interface. Sclerotomic cells then infiltrate these cavities. Dorsal migration of sclerotomic cells also occurs during this period. These migrating cells settle in regions that ultimately will become the neural arches.13 This occurs in stages 10 through 12 of embryologic growth. Anterolateral movement of sclerotomal cells occurs in stage 13. These pockets provide primordium for future growth of the ribs. Stages 15 and 16 are characterized by fissuring and flexures of the notochord alongside vertebral body and intervertebral disc primordium.6
The period of cartilage formation in vertebral development takes place during embryonic stages 17 through 23. This chondrification begins in the vertebral bodies and subsequently migrates dorsally into the neural arches.6 During the latter part of stage 23 the dorsal aspect of the neural arches begins to deviate medially.
Bone formation starts in the early fetal stage of growth with ossification of the cervical and thoracic regions followed by lumbar and sacral vertebral segments. This final period continues on into infancy and adolescence.14
Growth of spinal nerves begins with formation of the spinal cord. This initial differentiation involves clumping of neural crest cells in embryologic stage 12.15 Over the next three stages of development the neural crest cells form spinal ganglia that expand and move ventrally. The first ventral root fibers occur during stage 13. The primordium of the cervical and brachial plexuses also forms during this stage. Stage 14 is marked by complete development of all motor roots from C1 to S2 and growth of intersegmental anastomoses between these motor roots.16 Shortly after this stage the motor nerve roots move away from the spinal ganglia and toward their respective intervertebral foramen. Formation of the spinal nerve by mingling of motor and sensory root fibers occurs in stage 15. The roots enter the medial aspect of the foramen during stage 18, and progress completely into the foramen by stage 23. Other motor nerve development worth noting are the following: at the end of stage 14 the motor nerve fibers from the L2–4 segments moving a short distance into the leg.17 Creation of the brachial plexus begins in stage 16 and progresses through stage 23. At this point the major motor subdivisions of the brachial plexus resemble those of the adult.15 Stage 16 also heralds formation of the lumbar plexus and the individual femoral, obturator, tibial, and peroneal nerves, and stage 17 marks development of the hypogastric, pudendal, genitofemoral, and ilioinguinal nerves.15
The sensory nerves develop in a similar manner as their neighboring motor fiber counterparts. As discussed above, the formation of sensory spinal ganglia anlage begins in stage 12 of embryologic development.18 Sensory root fibers grow out first centrally from the ganglia to make connections with the spinal cord in stage 14. In the following stage the fibers move peripherally to blend with the motor root fibers to form the spinal nerve. Entrance to the intervertebral foramen is gained by the end of embryologic stage 16, and complete access to the foramen by all sensory nerve roots is gained by stage 23.19
The autonomic system also begins its formation during the middle embryologic stages of development. Unlike the motor and sensory nerve roots, the autonomic nerve differentiation begins with primordium formation in the lower thoracic, esophageal, and upper abdominal regions rather than in the cephalocaudal growth of the vertebra and nerve roots.20 This occurs at embryologic stage 13. This primordium continues differentiation in the lower cervical to sacral region in stage 14. The first white communicantes fibers can be identified in the T2–9 level at this stage.15 Anlage of the autonomic pelvic plexus begins in stage 15.21 Stage 16 is marked by continued growth of the white rami communicantes through the L3 spinal level. The sympathetic chain also has a cephalad movement from the thoracic to upper cervical segments during this stage. The first gray rami communicantes begin to appear at the C8–T1 level around this time (days 37–42).22 Auerbach’s plexus begins differentiation in embryologic stage 17. Continued growth of gray rami communicantes occurs through stage 23 with formation of autonomic nerve fibers to the S5 spinal level.16
The development of the meninges originates from a thin cellular network that exists between the somites, notochord, and neural tube.6 Its early formation is termed the ‘meninx primitiva.’ It serves as a host site for the migration of cells and eventual development of the meninges. This meninx primitiva generation occurs in embryologic stage 15, and grows to surround the complete neural tube in stage 16. In the following stage, the denticulate ligaments begin to form. By stage 18 the spinal canal has started to take shape with cavitation of the meninx primitiva quickly following in embryologic stage 19. Evidence of pia mater structure formation also occurs during this time. It consists primarily of meninx primitiva cells with a small contribution from neural crest cells. Formation of the dura mater in the cervical and thoracic regions follows pia mater growth in stage 20. It consists of mesodermal cells from sclerotomal and meninx primitiva origins. It initiates growth anterior to the spinal cord with progression longitudinally and circumferentially around the cord. Dura mater growth reaches the lumbar region by stage 22, and completely surrounds the spinal cord end to end at stage 23. Construction of the epidural space also begin during this last stage. Arachnoid mater development continues to be poorly understood at this time. It is believed to be the last of the meninges to form. This occurs after completion of the embryologic stages, i.e. post 60 days fertilization.
Fetal development
All major structures of the spine are clearly distinguishable by the end of the embryologic period of spinal growth. The next stage of development occurs during the fetal period. Its temporal boundaries include the termination of stage 23 of the embryologic period to birth. As discussed above, chondrification of the vertebra begins in the embryological period. It can be identified in the pedicles, lamina, and transverse processes of vertebrae prior to the fetal period. Early into the fetal period these chondrification centers enlarge and migrate into the posterior arches and vertebral bodies. Fusion of the posterior vertebral structures and dorsal growth of the spinous processes occur at the end of the first trimester (12th week).23 This period is also marked by the onset of ossification anteriorly in the body and posteriorly in the lamina of the vertebra. These two bony growth centers progress in a dissimilar manner. Ossification of the vertebral body first occurs in the thoracic region and spreads bi-directionally towards the cervical and lumbar segments. The laminar bony growth takes place in a more typical cephalocaudal pattern from cervical to coccygeal segments.24 In addition, we see a longitudinal growth of the spinal structures during this fetal period. This growth is demonstrated by measurement of the vertex-coccyx segment over this period. At 3 months, this segment measures about 10 cm long, and reaches to roughly 34 cm at birth.25
The intervertebral disc undergoes changes during this fetal period. The end of the embryologic stage and beginning of the fetal period see the notochord expand into the center of the disc. This cell in-growth to the disc forms a strandlike appearance that is implanted into an amorphous mucoid substance called chorda reticulum.26 The embryologic cartilage surrounding the expanding notochordal cells begins to have collagen fibers formed within it around the 10th week of gestation.27 Interestingly, their growth patterns are identical to those in the adult.28 Their formation is well established by the 6th month of fetal growth. In addition, the neighboring anterior and posterior longitudinal ligaments begin to develop around the same time as the annular fibers. They grow from perivertebral mesenchymal cells.29 As the fetal period continues, the chorda reticulum enlarges and radially expands. During the terminal stage of this period the chorda reticulum cells bordering the annular cartilage themselves begin a transition to fibrocartilage cells. These cells arrange themselves into patterns similar to those seen in the collagen fibers of the adult anulus fibrosus.26,30
Postnatal development
This final stage of development takes place from birth to adulthood. It is by far the longest of the three stages in duration. In contrast, this stage involves minimal plastic changes in the spinal tissues. This period primarily involves tissue maturation and longitudinal growth of the spine.25 The spinal maturation process involves continued ossification of the vertebrae. This is a continuation from its onset in the fetal period. As discussed above, the vertebrae typically have three centers of ossification, one in the ventral vertebral body and one in each of the lamina. At birth, approximately 30% of the spine is ossified, primarily from these ossification centers. The articular processes, transverse processes, spinous processes, and the last few sacral and coccygeal segments still demonstrate a cartilaginous pattern at birth.31 Hyaline cartilage also still persists in the superior, inferior, and peripheral portions of the vertebral body. A striking feature at birth is that all pars interarticularis of the cervical, thoracic, and lumbar spine are ossified at birth. Failure of this pars ossification at birth has clinical implications for development of spondylolysis and spondylolisthesis in adulthood.32
Except for the first two cervical vertebrae, the growth and development of the cervical spine is fairly uniform.33 The atlas, like other cervical vertebrae, has three centers of ossification. Unlike other cervical vertebrae, the atlas does not possess a body or centrum. In this case, the anterior ossification center forms in the anterior arch of the atlas. This anterior ossification center is visible on radiography in about 20% of neonates.34 By the end of the first year the anterior center is readily apparent on radiography. The neural arches ossify at 6–12 months of age.35,36 The posterior arches typically fuse around 3 years of age. Prior to this fusion, a secondary ossification center may be seen in the posterior tubercle.33 Other potential secondary ossification centers include the anterior tubercle and at the tip of the transverse processes. The posterior arches may fail to fuse, leaving a bifid process in place. This bifid process does not hold any potential pathologic consequences to the spine. It should typically be regarded as a normal variant similar to spina bifida occulta of the S1 posterior arches.
The axis has five primary and two secondary centers of ossification. Three of the primary centers are typical placed in the centrum and the bilateral posterior arches. These posterior arches normally fuse together between 2 and 3 years of age.31 The remaining two primary ossification centers appear during the 20th–24th weeks of gestation. They appear at the base of the cartilaginous rostral projection at the anterosuperior aspect of the axis. A secondary ossification center occurs in the apical portion of the dens around the end of the second year of life. This apical ossification center is phyletically derived from the centrum of the first cervical vertebra. The secondary center in combination with the two primary centers at the base of the odontoid result in a persistent cartilaginous region in the central area of the odontoid. This area is homologous for the intervertebral disc between the atlas and axis.35 The last secondary ossification center forms in this cartilaginous area in the late teens and usually completes union of the apex and base of the odontoid in the mid twenties. Absence of this last ossification step results in failure of fusion of the apical portion of the odontoid to the base of the axis. This results in a condition called os odontoideum. The presence of this condition can be easily confirmed by magnetic resonance imaging. The stability of the atlantoaxial joint then falls directly on the atlantoaxial ligaments and the degree of fibrous union between the dens and the body of the axis.35 Partial ossification of this site can occur, which can create the appearance of a fracture through the middle to inferior aspect of the dens. The rate of bony growth can also be very slow at this site, causing the area to fuse late in life.
The remaining cervical segments all develop in a more traditional pattern to the thoracic and lumbar segments. As noted above, they have three primary ossification centers: one located in the vertebral body, and one in each lamina. The vertebral body is almost entirely ossified by 6 years of age, and the neural arches approximate posteriorly at around 2–3 years of age.31 The epiphyseal plates of the vertebral bodies do not undergo ossification until completion of growth. This occurs around the twentieth year of life. This bony growth progresses in a radial pattern in a peripheral to central direction. However, this inward growth is arrested to leave a central cartilaginous portion that becomes the endplate zone in the adult. The pedicles remain cartilaginous until the sixth year of life when their ossification process is complete and they fuse with their respective vertebral bodies.37 Secondary ossification sites occur at the transverse processes and spinous processes around the fifteenth to sixteenth year of life. These areas usually fuse in the second to third decade of life. An accessory secondary ossification site may occur in the transverse process of the seventh cervical vertebrae.32 If this accessory center fails to fuse with the main secondary center in the transverse process, the accessory site may progress to formation of a cervical rib.33
The intervertebral disc also undergoes changes during the postnatal period. There is a progressive decline in the notochordal mesenchymal cells at birth. This is accompanied by a steady increase in the mucoid substance of the nucleus pulposus. A fibrocartilage capsule surrounds these diminishing notochordal cells and increasing mucoid substance. This change is typically evident by the 5th month of growth.38 By 4 years of age there are no active notochordal cells within the nucleus.39 After this point, the nucleus pulposus halts any further growth. Future growth of the disc thereafter occurs through the anulus fibrosus. This growth primarily involves production of fibrous elements. As years advance there is in-growth of fibrous tissue into the nucleus pulposus with resultant progressive loss of its liquid content.40 The intervertebral disc in the newborn has a relatively wedge-shaped appearance with the nucleus located posteriorly in the disc.41 The nucleus reverses position and is primarily anteriorly placed by 2 years of age.41 As the child assumes a more upright posture and gait, the nucleus takes on a more central position in the disc. This occurs around the fourth to the eighth year of life.30,41 A significant horizontal and longitudinal growth phase occurs in the disc after birth. This growth progresses late into adolescence.
A more generalized discussion on growth after birth and to adulthood will be summarized to demonstrate the developmental requirements of the spine. It is marked by two main growth spurts: one occurring after birth and the second during puberty.42 The early growth spurt takes place from birth through the age of five. Almost half of this initial growth occurs during the first year of life.41 As mentioned above, at birth the sitting height measures around 35 cm. After the first year of growth this sitting height increases to about 47 cm.25 In the following four years of growth, the sitting height increases from 47 cm to 62 cm, so that by the time the normal infant reaches 5 years age it has increased its height 27 cm. Over the next 5 years the truncal growth slows dramatically in comparison to the prior 5 years. It increases about 10 cm over this period. The pubertal, or adolescent, phase marks the last stage of linear spinal growth. This commonly occurs in the tenth through eighteenth years of life. During this final stage the second growth spurt occurs. Its onset is slightly different for each gender. In females it usually begins slightly earlier than in males. Most commonly, it occurs around the age of 11. There is an initial period of rapid growth over the first 2 years that is followed by a slower growth phase over the remainder of the pubertal stage. The early phase increases sitting height approximately 7 cm, while the slower second stage increases height by another 5 cm. In males, the pubertal stage usually has its onset around age 13. The male growth pattern is similar to that of the females. However, there is a slight increase in height versus the female growth during the first phase of this growth spurt.41
By the time we progress from birth to adulthood the spine has nearly tripled in size. This longitudinal growth is accomplished primarily through growth of the vertebral bodies. As discussed earlier, this growth occurs at the superior and inferior surfaces of the bodies.42 These regions persist as cartilaginous zones for continued growth and ossification of the vertebral body.43 The cartilage cells in these regions are arranged in a perpendicular-like pattern to the body. Thus, the longitudinal growth of the vertebral body is much the same as the growth of the metaphyses of long bones in the human body.25,44 Whereas cells closest to the vertebral body undergo ossification and unite with the vertebral body, other cartilaginous cells move toward the vertebral body and replace these ossifying cells.45 Hence, the vertebral body continues its longitudinal growth through this ossifying process.44 This growth terminates between the ages of 18 and 25.46 The growth plate then thins and calcifies. However, there remains a fibro-cartilaginous region neighboring the disc. This area becomes the adult vertebral endplate. Each level (i.e. cervical, thoracic, lumbar, and sacral) increases its vertebral height to different degrees. The average cervical and thoracic vertebral body height triples from birth to adulthood.23 The lumbar vertebral segments quadruple their longitudinal size by adulthood.23 At birth, the vertical length of the cervical spine measures about 3.7 cm. It doubles in length by 6 years of growth, during the first growth spurt, and nearly doubles its length again at the pubertal growth spurt. By the end of the pubertal growth spurt the cervical spine constitutes about 22% of the total spine length, and 15–16% of sitting height.23 In addition to vertical growth, the vertebral body also expands in a radial pattern.45 Periosteal ossification is primarily responsible for this horizontal growth. Most expansion, similar to vertical growth, occurs during the two main growth spurts.47
FUNCTIONAL ANATOMY
Bony structures
The bony structures of the cervical spine consist of seven vertebrae connected together by a system of complex articulations that primarily serves four main functions: (1) to support the head, (2) to allow for an extensive, precisely controlled range of motion for the head, (3) to provide a protective covering for the spinal cord and nerve roots, and (4) to supply an entrance and exit zone for neurovascular structures. The cervical spine is unique, as the morphology of the two cephalad segments is markedly different from the rest of the cervical spine.35 We will first discuss the bony anatomy of the atlas and axis, followed by the third through seventh cervical segments.
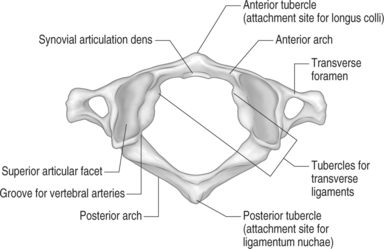
The atlas is the first cervical vertebrae. Along with the axis, it serves to connect the cranium to the spine. Metaphorically speaking, it acts as the ‘cradle’ for the occiput. It is unique to the other cervical segments because it lacks a vertebral body. In place of the body the atlas develops an anterior arch that runs almost perpendicular to the connecting lateral masses. A small tubercle forms in the midline on the anterior aspect of the arch. This tubercle serves as the insertion point of the longus colli muscle.48 A rounded cavity can be seen on the posterior surface of the anterior arch. This depression marks the synovial articulation of the odontoid process. The paired, cylindrical lateral masses contribute their articular processes, or zygapophyseal joint, to the atlanto-occipital and atlantoaxial articular joints. The underside of the lateral masses are slightly concave, directed medially and posteriorly to articulate with the superior articular process of the axis. The cephalad portion of the lateral mass is oriented superiorly and medially to interdigitate with the articulation of the occipital condyles. Small tubercles can be noted on the ventromedial aspect of the pillars. These tubercles serve as the attachment sites of the transverse atlantal ligaments.35 These ligaments hold the odontoid against its anterior synovial membrane. The ring-shaped atlas is closed dorsally by the formation of a posterior arch connecting to each lateral mass. At the juncture of the lateral masses to the posterior arch there is a small groove that marks the passing of the vertebral arteries towards the foramen magnum.49 The transverse processes of the atlas are short and compact structures. Within each transverse process exists a small rounded transverse foramen for passage of the vertebral artery.49 Unlike other cervical segments, the atlas does not have a spinous process. There exists a small tubercle lying in the midline on the dorsal aspect of the posterior arch.33 This tubercle serves as the attachment site of the ligamentum nuchae.35 As mentioned above, the lateral masses of the atlas combine with the occiput to form the occipital–atlantal (OA) and the axis to form the atlantal–axial (AA) articulations (Fig. 46.1).50 The primary motion of the OA joint is flexion–extension moments.51 There is minimal appreciable axial rotation of the OA joint. The reason for this limitation is secondary to the concavity of the superior portion of the lateral masses. The concavity essentially encloses the condyles within its bony walls, preventing the required anterior and posterior translation for rotation to occur.52 However, the cupping nature of the superior lateral masses does allow for a sagittal rotation (i.e. rolling) movement to occur through the OA joint. This nodding motion is achieved through multiple actions occurring through the OA joint. As the head nods forward the condyles roll forward in the sockets of the atlas. Concomitantly, the condyle slides up the concave anterior wall of each socket. Posteriorly and inferiorly directed forces by the flexor muscles, weight of the head, and tension of the OA capsule prevent anterior dislocation of the condyles and maintain them snugly within the lateral mass sockets.52 The reverse process occurs in extension. Normal flexion and extension of the AO joint has a range of 14–35 degrees.53–55
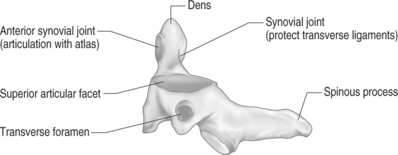
The second cervical vertebra is commonly referred to as the axis (Figs 46.2. 46.3). This name is very proper for this cervical segment as its main function is to allow the atlas to rotate upon it.48 Like the atlas, it also serves to transmit the load of the head to the remainder of the cervical spine. It is regarded as the strongest of all the cervical vertebrae. Its body and posterior arch are similar to those of the lower cervical segments. The uniqueness of the axis is the presence of the odontoid process. It is a vertical cylindrical bony growth that emanates from the superoventral aspect of the axis’s body. The tip of the dens (odontoid) typically terminates at the superior border of the anterior arch where it forms an articulation. A small, smooth, oval indentation is present on the base of the dorsal aspect of the dens. This area is occupied by a synovial joint that serves to protect the transverse ligament from contact with the bony dens.33 A synovial articular process is also present on the ventrosuperior aspect of the dens. It is commonly referred to as the median atlantoaxial joint.52 The dens acts as a pivot for the atlas directly above to spin and glide around to achieve axial rotation. It forms an articulation with the dorsal aspect of the atlas’s anterior arch. A large bifid spinous process is usually present at this level. This spinous process is the first one to be palpated in the superoposterior groove of the neck.48 The inferior articular process of the lateral masses is directed caudally. It forms a facet joint with the superior articulating process of the third cervical vertebrae. A convex and medial-to-lateral orientation is present in the superior articulating process of the lateral masses. This orientation guarantees that the downward force of the atlas is transmitted directly through the central portions of the lateral masses of the axis.35 The primary motion of the atlantoaxial (AA) joint is axial rotation. This requires anterior displacement of one lateral mass with concomitant posterior translation of the opposite lateral mass.52 The alar ligaments are the primary structures to restrain axial rotation. The lateral AA joint capsules are thought to play a minor role in limiting rotation.56,57 The normal range of axial rotation is 0–45 degrees in each direction.58 It accounts for 50% of the total rotation of the neck, and gives almost all rotation over the first 45 degrees. Flexion and extension also occur at the AA joint. Flexion occurs by anterior translation of the atlas on the axis. This causes separation of the anterior arch of the atlas from the dens. Anterior translation is limited by increased tension of the transverse and alar ligaments.56,59 Extension occurs as a result of a slight posterior bend in the dens. This allows the anterior arch of the atlas to move dorsally and superiorly on the dens. Extension is limited by the anterior arch contacting the odontoid. The total range of motion of the flexion–extension moment is approximately 10 degrees.60
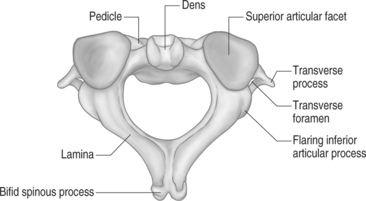
The remaining cervical segments all demonstrate a similar bony appearance (Figs. 46.4, 46.5). In general, these vertebrae are required to bear less weight than the thoracic and lumbar segments so they are smaller and thinner than their caudal counterparts. The conventional cervical vertebrae consist of two main parts: the cylindrical, blocklike ventral vertebral body, and the dorsal vertebral arch. This posterior arch is formed by a number of bony components: the pedicles, lamina, spinous process, transverse process, and superior/inferior articulating processes. Their fusion creates vertebral foramens at each cervical segment that serve to create a longitudinal spinal canal for housing of the spinal cord and nerve roots.
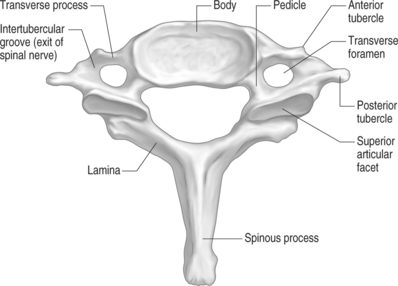
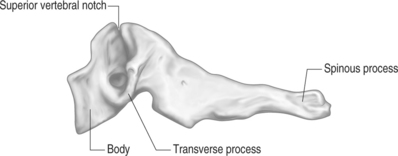
The pedicles are short, cylindrical bony structures projecting from the superior and dorsolateral aspect of the vertebral bodies. Connecting to each pedicle is a thin, flat, platelike bone called the lamina. These bony plates project dorsomedially until they fuse together in the sagittal plane of the body. At the dorsal juncture of the lamina projects a small, flat, rectangular bone called the spinous process. This bone typically has a bifid appearance at the second through sixth cervical segments. The seventh cervical spinous process tends to be very long. Hence, it has been termed the vertebra prominens, and is the most palpable of the cervical vertebrae. The transverse processes project laterally from the junction of the pedicle and lamina.32 The cervical spine transverse processes are rather short in comparison to those of the thoracic and lumbar regions. In addition, these processes also have a transverse foramen just lateral to the pedicle–transverse process junction. Through these foramens pass the vertebral artery on its way toward the foramen magnum. The exception to this rule is the C7 transverse foramen, which only transmits veins.49 A more concise description of this artery will be discussed later. There are two tubercles located on the lateral edge of the transverse process. One is positioned anteriorly and posteriorly. Each tubercle serves as insertion points for the ventral and dorsal cervical musculature. An intertubercular groove exists proximally on the transverse process. This groove has important clinical implications, as this is where the spinal nerves exit the foramen. At this position, the nerves pass just dorsal to the vertebral artery as it passes through the neighboring transverse foramen.49 The superior and inferior articular processes project vertically at the juncture of the pedicle and lamina. In general, the superior articular processes face dorsally and cephalad at about a 45 degree angle to the transverse plane.51 This superior position permits the articular process to bear the weight of the pillar above.52 The height of the superior articular process increases in a cephalocaudal direction. It plays a major role in controlling cervical spine segmental motion in the sagittal plane. As a general rule, the smaller the superior articular process, the greater the segmental flexion.51 In other words, there is a much greater range of sagittal motion seen through the upper cervical segments compared to the lower segments due to this phenomenon. The inferior articular processes mirror the superior articular process position by facing ventrally and caudally. The approximation of a superior articular process from a vertebra with an inferior articular process to the vertebrae above results in the formation of a facet, or zygapophyseal, joint.50
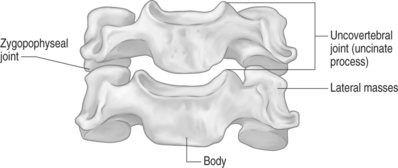
The uncinate processes are paired bony protuberances that project superiorly from the posterolateral aspect of the superior endplate of the C3 through C7 vertebral bodies (Fig. 46.6). It is believed they represent phylogenetic remnants of ancient costovertebral joints in birds and reptiles. These processes are absent in infants and toddlers. They develop in late childhood and early adolescence. Their function is unknown. However, some investigators believe that they function to bolster the cervical disc laterally to prevent herniation.61 Other studies suggest these uncinate processes act as guide rails for the translation which occurs in the cervical discs during flexion and extension.62 A cleft formed between this uncinate process and the medially located cervical disc creates the uncovertebral joint, or joint of Luschka.63 As these clefts do not contain articular cartilage, synovium, or a capsule they should not be referred to as a joint. These clefts allow for a large degree of movement between the vertebral bodies and the intervertebral disc,64,65 particularly in axial rotation.66






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


