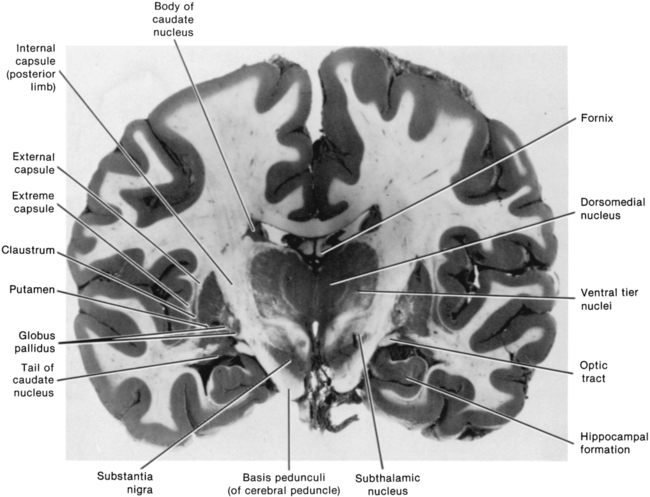
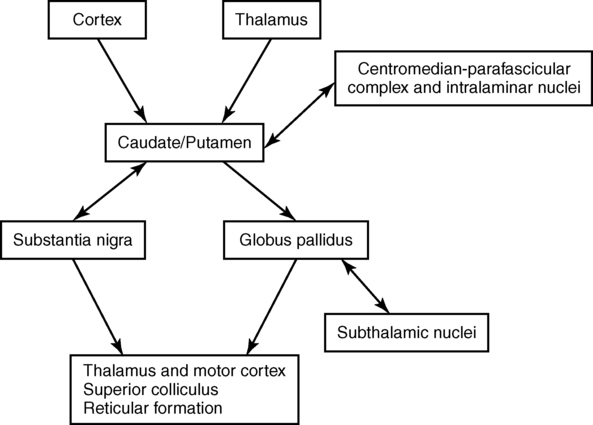
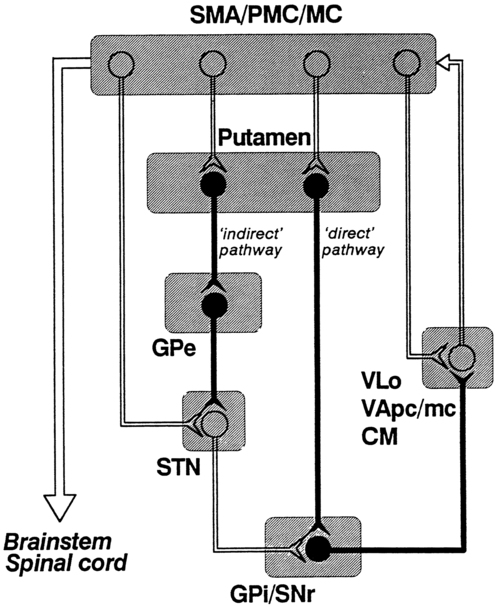
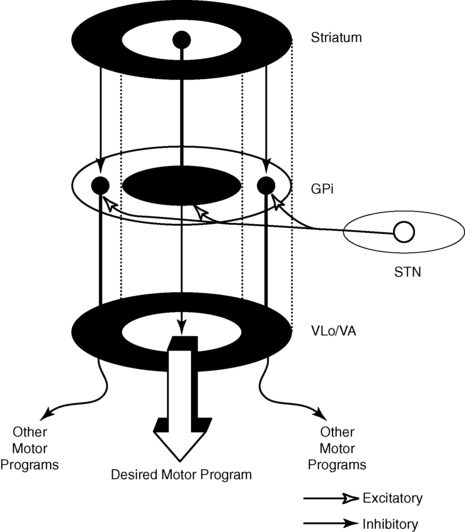
After reading this chapter the student or therapist will be able to: 1. Describe the circuitry of the basal ganglia. 2. Relate the anatomy and physiology of the basal ganglia to its roles in sensorimotor and cognitive processes. 3. Use the information on anatomy, physiology, and pharmacology to explain the signs and symptoms seen in classic disease states—for example, Parkinson disease, Huntington disease, and dystonia. 4. Develop an evaluation plan for patients with diseases of the basal ganglia. 5. Develop an intervention plan for patients, with the rationale for treatment methods. 6. Determine treatment effectiveness, especially in the case of degenerative disease. 7. Integrate the information in this chapter with the information provided in Section I of this book to develop treatment plans for patients with metabolic or toxic disorders. This chapter considers the degenerative, metabolic, hereditary, and genetic disorders that typically have their onset in adulthood, including Parkinson disease, parkinsonian syndromes, Huntington chorea, Wilson disease, dystonias, heavy metal poisoning, and drug intoxication. Because of the wide variety of diseases with their wide variety of causes, the concentration is on understanding the clinical problems and commonalities that exist within this grouping. In general, the practice parameter of the diseases discussed in this chapter is the physical therapy diagnostic parameter 5E: Impaired motor and sensory integrity associated with progressive disorders of the central nervous system, from the Guide to Physical Therapist Practice.1 Although the occupational therapy guide does not classify practice parameters in that manner, the concepts and clinical reasoning process can be used by both professionals. The predominant area of the brain affected by these disorders is the basal ganglia: this group of central nervous system (CNS) structures is therefore discussed in some detail. The most commonly seen disorders affecting the basal ganglia include Parkinson disease, Huntington chorea, and dystonias, including drug-induced dyskinesias. All of these medical diagnoses involve impairments in muscle tone, movement coordination and motor control, and postural stability and the presence of extraneous movement. Taken together, these disorders now affect approximately 1 million people in the United States.2–4 The dorsal or sensorimotor basal ganglia are composed of three nuclei located at the base of the cerebral cortex—hence their name. These nuclei are the caudate nucleus, the putamen, and the globus pallidus. Two brain stem nuclei, the substantia nigra and the subthalamic nucleus, are included as part of the basal ganglia because they have a close functional relation to the forebrain nuclei. In addition, connections between the basal ganglia and the pedunculopontine nucleus (PPN) are important in regulating underlying tone. Other parts of the basal ganglia, the ventral basal ganglia, are intimately related to the limbic system and are discussed in Chapter 5. The anatomical location of the various parts of the basal ganglia is shown in Figure 20-1. Functionally, the basal ganglia can be divided into an afferent portion and an efferent portion (Figure 20-2). The afferent structures are the caudate and putamen. They receive input from the entire cerebral cortex, the intralaminar thalamic nuclei, and the centromedian-parafascicular complex of the thalamus as well as from the substantia nigra and the dorsal raphe nucleus, both located within the brain stem. The projections from the cortex are systematically arranged so that the frontal cortex projects to the head of the caudate and putamen and the visual cortex projects to the tail. In addition, the prefrontal cortex projects mainly to the caudate, whereas the sensorimotor cortex projects mainly to the putamen.5–8 Projections from the cortical regions that represent the proximal musculature, and those from the premotor regions may be bilateral.6,9–11 These close and profuse connections between the cortex and the basal ganglia suggest a close interfunctional relationship. The projections from the thalamus to the caudate-putamen are also somatotopically arranged. The heaviest projections are from the centromedian nucleus, and these nuclei also receive massive input from the motor cortex.7–10 The somatotopic arrangement of the cortico-striatal–thalamic-cortical pathways is maintained throughout the loop. This finding has led to an important functional hypothesis that the basal ganglia form parallel pathways subserving specific sensorimotor and associative functions.5 The putamen is linked to the sensorimotor functions and the caudate to the associative, including cognitive functions.9,12 As knowledge of the circuitry of the basal ganglia has advanced, so has the knowledge regarding the microscopic structure. The caudate-putamen looks somewhat homogeneous because of the predominance of one cell type. Careful analysis using precise staining methods has demonstrated the appearance of patches within these nuclei. It is hypothesized that this organization is important for the ability of the basal ganglia to modulate ongoing sensory input and choose the appropriate motor response.12 The intrinsic structure of the caudate-putamen also suggests that at least nigral input occurs in a way that could immediately modulate the input coming from the cortex.13,14 The input that has been processed in the caudate-putamen is sent to the globus pallidus (pallidum) and substantia nigra (nigra), which constitute the efferent portion of the basal ganglia. The globus pallidus and substantia nigra are each divided into two regions. The globus pallidus has an external and an internal region; the substantia nigra consists of the dorsal pars compacta and the ventral pars reticulata. Embryologically and microscopically, the internal segment of the globus pallidus and the pars reticulata of the substantia nigra are similar. These two regions are the primary efferent structures for the basal ganglia. The projections from the caudate and putamen to the pallidum and nigra maintain a somatotopic arrangement.10,15,16 From these structures the information is transmitted to the thalamus and then to the cortex, still maintaining somatotopy. The superior colliculus, the PPN, and other, less defined brain stem structures (perhaps the reticular formation) also receive pallidal and nigral output. All output of the basal ganglia has then been processed through the globus pallidus and/or the substantia nigra before proceeding to other areas of the brain (see Figure 20-2). Information processed in the basal ganglia can influence the motor system in several ways, but no direct pathway to the alpha or gamma motor neurons of the spinal cord exists. The first route is the projection to the ventroanterior and ventrolateral nuclei of the thalamus, which then project predominantly to the premotor cortex. Another pathway is through the superior colliculus and then to the tectospinal tract. Pathways exist from the globus pallidus and substantia nigra that terminate in areas of the reticular formation (e.g., the PPN) and thus may influence the motor system through the reticulospinal pathways. Anatomically the basal ganglia are therefore in good position to affect the motor system at many levels. Many of these connections are also areas that receive cerebellar input, and thus these two regions of the brain have ample opportunity to further integrate movement responses.17 The basic circuitry of the basal ganglia comprises two loops.7 The loops for the sensorimotor system are shown in Figure 20-3. The direct loop is the loop that begins in the motor regions of the cortex and projects to the putamen and then directly to the globus pallidus, the internal segment, and on to the thalamus. The indirect pathway adds the subthalamic nucleus between the globus pallidus, external segment, and internal segment before sending the signal on to the thalamus. The subthalamic nucleus also receives direct input from the premotor and motor cortex as well as from the pallidum.18,19 The darkened neurons represent inhibitory connections, and the open neurons represent excitatory connections. In general, the direct pathway, by disinhibition, activates the thalamocortical pathway; the indirect pathway inhibits the thalamocortical system. The role of these loops in normal and diseased states is clarified in the discussion of the physiology and pharmacology of the basal ganglia. The caudate and putamen are composed of neurons that fire slowly; the globus pallidus neurons fire tonically at high rates. The low firing rates of the caudate-putamen are partially a result of the nature of thalamic inputs. Input from the cortex seems to have priority over input from the thalamus and substantia nigra. These data indicate that the cortex is instrumental in regulating the responsiveness of caudate and putamen neurons.20 In turn, basal ganglia stimulation may prepare the cortex for subsequent inputs; this might be especially important when a response must be withheld until an appropriate stimulus occurs, such as keeping the foot on the brake until the light turns green.20–23 Mink hypothesized that basal ganglia inputs to the cortex activate only the most necessary pathways and inhibit all unnecessary pathways (Figure 20-4).24 The pattern of neuronal firing in the direct and indirect pathways also suggests that the basal ganglia modify input to the cortex. The neurons of the efferent portion of the basal ganglia respond with either phasic increases or phasic decreases in activity, which in turn will affect the activity in the thalamus and hence the cortex. A decrease in activity of the internal segment of the globus pallidus removes inhibition to the thalamus and thus enables cortical activation. Whether the two pathways are activated concurrently or whether different activities activate the two pathways separately is not yet known; either way, the basal ganglia would have a role in cortical activation and modulation. One of the current views in relationship to disease processes is that an underactive direct pathway and/or an overactive indirect pathway would lead to decreased activation of the cortex and hence bradykinesia and akinesia, whereas an overactive direct pathway and/or underactive indirect pathway would lead to the presence of extraneous movements (see Figure 20-3).6,25 How do these pathways relate to everyday function? Rigidity could be explained by too much muscle activity (through the pathways from the basal ganglia to the PPN and on to the spinal cord). Akinesia and bradykinesia typical of individuals with Parkinson disease are caused by insufficient excitation or too many conflicting patterns of movement. Increased extraneous movements are characteristic of basal ganglia diseases and can be attributed to the dysfunctions within these pathways. If the amount of muscle activity and the sequence and timing of activation are inappropriate, the individual will have difficulty in selecting the environmentally appropriate behavior.24–27 Aldridge and colleagues found that the basal ganglia were modulated dependent on the purpose of the impending movement.27 The earliest view of the basal ganglia came from Willis in 1664. He hypothesized that the corpus striatum received “the notion of spontaneous localized movements in ascending tracts . . . . Conversely, from here tendencies are dispatched to enact notions without reflection [automatic movements] over descending pathways” (p. 7).28 Willis possessed great insights in the discussion of the signs and symptoms of basal ganglia disease. Magendie in 1841 demonstrated that removal of the striatum bilaterally produced compulsive movements, whereas removal of only one striatum produced no visible effect.29 Studies by Nothnagel30 demonstrated that lesions of the nigra tended to produce immobility. With the advent of the use of electrical stimulation in the late nineteenth century, further information on the function of the basal ganglia was gathered. Stimulation of the caudate nucleus did not (and does not) produce movement of muscles or limbs as occurs with stimulation of the motor cortex. However, at higher levels of current, total body patterns and postures were usually evoked. The earliest stimulation of the caudate nucleus produced an increase of flexion of the head, trunk, and limbs and tonic contraction of the facial muscles.31 These early studies are mentioned because of the insights they provide for the symptoms of the disorders of today. Contemporary experiments using lesion paradigms show a wide variety of motor problems in a variety of animals. Hypokinesia, a decrease or poverty of movements, a decreased amount of exploration of novel environments, and a tendency to assume a fixed posture are the most common problems after a lesion in the basal ganglia. These motoric dysfunctions are seen regardless of the method by which the lesion is made: pharmacologic, surgical, or by stimulation. In essence, movements are altered in scale (related to the gain), take longer for completion, and take place under altered conditions of antagonistic muscle interactions (e.g., contraction).32–40 The hypothesis that the basal ganglia are involved in movement initiation and preparation is an area of some research disagreement. A “readiness potential,” recorded from the scalp of human beings before movement and thought to reflect basal ganglia activity, is more apparent in complex than in simple movements, for example, before dorsiflexion with gait but not before dorsiflexion when sitting.41–44 Neuronal recordings from awake, behaving animals found that units in the basal ganglia alter their activity before changes in the electromyographic activity of the prime movers of the task.45–51 Studies recording from multiple units in animals moving freely in their home environment suggest that neurons in the caudate-putamen and in the substantia nigra are activated in sequential, purposeful movements.27 The basal ganglia have been implicated in the process of posture and postural adjustments. People with diseases of the basal ganglia assume flexed or other fixed postures as the disease progresses (Figure 20-5). In addition, these individuals have decreased postural stability and are therefore at risk for falls. Animal experiments indicate that a deficit exists in determining response based on one’s own body position, or “egocentric localization.”52–54 This deficit decreases the ability of a person with basal ganglia disease to modify a postural response to the precise environmental demands. Martin,55 in his extensive studies of individuals with Parkinson disease, was the first to describe severe disturbances in posture, especially when vision was occluded. Melnick and colleagues56 showed that a decrease in static postural adjustments in persons with Parkinson disease could be seen early in the disease process.57 Bloem and colleagues58–60 and Visser and colleagues61,62 meticulously studied the reflexes involved in postural adjustments and described deficits in the longer loop reflexes but not in the short latency reflex associated with the stretch reflex. Others have investigated the interactions of the sensory systems involved in balance in those with Parkinson disease.62–66 Bloem and colleagues60 and Visser and colleagues62 concluded that postural instability was caused by a decrease in proprioception. In a recent review of proprioception and postural stability and motor control, Nicola and colleagues also describe the kinesthetic and proprioceptive deficits in people with Parkinson disease. Nicola and colleagues concluded that there was a “failure” in the body map similar to the failure in egocentric localization described previously.54 A decrease in the ability to use proprioceptive and kinesthetic information to properly scale the input and response also contributes to a loss of balance reactions. The basal ganglia are not solely motor systems. The previous paragraphs demonstrate the role of the basal ganglia in sensory integration. The basal ganglia are also involved in cognitive functions and responses associated with reward.36,37,48,50,67–70 Researchers have found that learned movements are more affected by basal ganglia lesions than reflexes, that neurons in the basal ganglia are responsive to some sensory input, especially proprioceptive input, and that neurons in other parts of the basal ganglia are responsive to reward and anticipation of the reward.26,71,72 Klockgether and Dichgans73 as well as Jobst and colleagues74 found that patients with Parkinson disease likewise had impairments in kinesthesia and that as a person moved a limb further from the body’s center, kinesthetic sense decreased. Schneider and colleagues75 found that animals that developed parkinsonian symptoms from a neurotoxin had deficits in operantly conditioned behavior. They suggested that the decrease in performance resulted from a “defect in the linkage” between a stimulus and the motor output centers. These sensory difficulties may be important factors in evaluation and treatment of basal ganglia diseases, especially those associated with dystonia. The basal ganglia appear to be involved in the process of withholding a response until it is appropriate.76 A deficit in alternation of response may be the result of a tendency toward perseveration of a previously reinforced cue.77 Additional deficits exist in remembering or relearning tasks requiring a temporal sequence.78 Graybiel26 integrated the behavioral findings with information from her anatomical and chemical studies to suggest that the basal ganglia are important in providing behavioral flexibility. She hypothesizes that the basal ganglia are involved in procedural learning that leads to the development of habits. These habits become routine and are easily performed without conscious effort. Because these activities can proceed without thought, we are free to react to new events in our environment and to think. She and colleagues have performed electrophysiological experiments that explain this learning process, and these studies demonstrate great plasticity in basal ganglia networks.79 This enables the individual to select the proper movements in the proper environmental context. An elegant study by Brown and colleagues80 demonstrates a model of the basal ganglia that can reflect these cognitive and learning activities. Their model seems to integrate many of the functions of the basal ganglia with the physiology and pharmacology of the entire system. These cognitive dimensions are important to remember when developing a plan of care for a patient with basal ganglia dysfunction. Humans with basal ganglia disease also show problems in perceptual abilities, including deficits in tasks that involve perception of interpersonal and intrapersonal space.81 In pursuit-tracking tests individuals with Parkinson disease had particular difficulties in correcting errors77; if the motor system is inflexibly set, corrections can be made only by a complete reprogramming. The ability to perform cognitive activities involves integrating sensory information and, on the basis of this information, making an appropriate response. The basal ganglia seem to have a sensory integrative function as evidenced by experiments that show a multisensory and heterotopic convergence of somatic, visual, auditory, and vestibular stimuli.26,71,72 Segundo and Machne82 hypothesized that the function of the basal ganglia was not subjective recognition of the stimuli but rather in the regulation of posture and movements of the body in space and in the production of complex motor acts. Nicola and colleagues had similar conclusions.54 For movements to be properly controlled and properly sequenced, the two sides of the body need to be well integrated. There is anatomical evidence that suggests some means of bilateral control for the basal ganglia. A lesion of one caudate nucleus or nigrostriatal pathway produces a change in the unit activity of the remaining caudate.78,83 Studies of the dopaminergic pathway also indicate interactions between the two sides of the body.83 For this reason one may find deficits in function even on the “uninvolved” side of an individual with disease of the basal ganglia. It is also possible that diseases of the basal ganglia may go unnoticed until damage is found bilaterally. This summary of experimental results on the function of the basal ganglia illustrates several points. At least in some general way the basal ganglia are involved in the processes of movement related to preparing the organism for future motion and future reward. This may include preparing the cortex for approximate time activation, setting the postural reflexes or the gamma motor neuron system, organizing sensory input to produce a motor response in an appropriate environmental context, and inhibiting all unnecessary motor activity. Because of the multilevel involvement of the basal ganglia in movement, it is crucial that clinicians carefully observe all aspects of movement (simple and complex) with and without interference of sensory cues or performance of dual tasks as well as postural tone during examination and treatment and the responses to treatment (see Chapter 9). Before a detailed analysis of the diseases of the basal ganglia can be considered, a brief description of the neurotransmitters of this region is necessary. The most prevalent diseases discussed in this chapter indicate a deficit in specific neurotransmitters. The pharmacological treatment of Parkinson disease and, in the future, perhaps other “basal ganglia plus” diseases, is based on these neurochemical deficits. The basal ganglia possess high concentrations of many of the suspected neurotransmitters: dopamine (DA), acetylcholine (ACh), γ-aminobutyric acid (GABA), substance P, and the enkephalins and endorphins. This discussion, however, includes only the first three neurotransmitters. A diagram of the basal ganglia pathways, which includes the neurotransmitters, is shown in Figure 20-6. DA is the major neurotransmitter of the nigrostriatal pathway. It is produced in the pars compacta of the substantia nigra. The axon terminals of these dopaminergic neurons are located in the caudate nucleus and putamen. DA appears to be excitatory to the neurons in the direct pathway (GABA and substance P neurons) and inhibitory to the neurons in the indirect pathway (GABA and enkephalin neurons).2 This dual effect means that a loss of DA will lead to a loss of excitation in the direct pathway and an excess of excitation of the indirect pathway, leading to a powerful decrease in activation of the thalamocortical pathway. Several DA receptors exist; however, their chemical interactions permit the continued use of D1 and D2 receptor classes.7 The role of DA may modulate the effects of other neurotransmitters such as glutamate. Many new drugs (called the dopamine agonists) influence only one of these receptors. Recent experiments have been trying to determine which behaviors are mediated by which DA receptor in the hope that this research may lead to more effective drug treatment with fewer side effects. Because various drugs and chemicals can act as agonists (similar to) and antagonists (blocking the action of) of DA, they are used in treating disease involving the basal ganglia. Agonists include amantadine, apomorphine, and a class of drugs called the ergot alkaloids (e.g., bromocriptine). Amphetamine, which prevents the reuptake of DA, can enhance the effect of any DA present in the system. Antagonists include haloperidol, clozapine, and antipsychotic drugs of the phenothiazine class. With time these drugs may deplete the basal ganglia of DA and thus cause Parkinson disease or tardive dyskinesia. Similar effects on the DA system are observed in a single dose of methamphetamine (see Chapter 36).84 ACh is believed to be the neurotransmitter of the small interneurons of the caudate and putamen. It is presumed to inhibit the action of DA in this region and classically must be “in balance” with DA (and GABA). Dopaminergic axon terminals are found on cholinergic neurons. Substances that increase dopaminergic activity decrease release of ACh and vice versa.85 The antagonists of ACh, such as belladonna alkaloids and atropine-like drugs, were one of the first class of drugs used in the treatment of Parkinson disease. ACh antagonists are still used as adjuncts to treatment for patients with Parkinson disease. As some of the drugs to treat dementia are ACh agonists, care must be used when these are prescribed for the person with basal ganglia dysfunction, especially Parkinson disease. GABA is an inhibitory neurotransmitter that is found throughout the brain. In the basal ganglia it is synthesized in the caudate nucleus and putamen and transmitted to the globus pallidus and substantia nigra.86 GABA in the basal ganglia may permit movement to occur by allowing a distribution of neuronal firing. It also may provide a means of feedback inhibition in the efferent parts of the basal ganglia so that the program of activity is not repeated unless needed.86 Individuals with Huntington disease have a deficiency of this chemical. Although agonists of GABA exist (e.g., muscimol and imidazole acetic acid), a successful drug for the treatment of Huntington disease has not yet been found. This may be a result of either the ubiquitous nature of GABA or the very complex circuitry and interrelationships that exist among GABA, ACh, and DA. In addition to the transmitters discussed, co-transmitters may be found in the basal ganglia. Two such co-transmitters are cholecystokinin and neurotensin. The interactions of these co-transmitters may alter the sensitivity of DA receptors. Fuxe and colleagues87 suggest that the interactions of co-transmitters may alter the “set point” of transmission in synapses. They may therefore be important in one of the side effects of DA therapy, supersensitivity. Parkinson disease, first described by Parkinson in 1807, is a disease characterized by rigidity, bradykinesia (slow movement), micrography, masked face, postural abnormalities, and a resting tremor. As might be suspected from the review of functional physiology of the basal ganglia, the postural abnormalities include an assumption of a flexed posture, a lack of equilibrium reactions, especially of the labyrinthine equilibrium reactions, and a decrease in trunk rotation. Parkinson disease is among the most prevalent of all CNS degenerative diseases. Presently there are an estimated 1 million people in the United States with this disease, with approximately 60,000 new cases each year; the incidence is 4.5 to 20.5 and the prevalence is 31 to 347 per 100,000. (Refer to the list of websites at the end of this chapter.) Incidence increases with advancing age, and it is estimated that one in three adults over the age of 85 will have this disease.2 The personal and societal burden of Parkinson disease is great and includes the costs of actual treatment, the burden of caregiving, and the costs of lost earnings in patients under the age of 65.88 The cause of Parkinson disease remains unknown, and the consensus is that it is multifactorial.89,90 A slow viral process or long-term effects of early infection were implicated in postencephalitic parkinsonism. Some evidence indicates involvement of environmental factors and that interaction of environment and aging lead to a critical decrease in DA. Several investigators have found a link between growing up in a rural area and Parkinson disease; the important factors include pesticide use, insecticide use, and elements in well water.91–97 Accumulation of free radicals, cell death to excitatory neurons from toxins, and dysfunction of nigral mitochondria have all been implicated in the pathological process. The genetics of Parkinson disease is still debated. Although twin studies indicate that there may not be a single gene involved in Parkinson disease, as in Huntington disease, a family history may be an important risk factor.93,98–101 Very recently a large-scale study found two genetic loci to be associated with Parkinson disease.102 So the debate continues, with most neurologists agreeing that the multifactorial approach will yield the best opportunity to develop a cure. In view of possible treatment effects for Parkinson disease, it is interesting that a study by Sasco and others103 found an inverse relationship, albeit small, between participation in exercise or sports and later development of Parkinson disease. The loss of DA from the substantia nigra leads to alterations in both the direct and indirect pathways of the basal ganglia, resulting in a decrease in excitatory thalamic input to the cortex and perhaps a decrease in inhibitory surround that leads to the symptoms of Parkinson disease. Bradykinesia (a decrease in motion) and akinesia (a lack of motion) are characterized by an inability to initiate and perform purposeful movements. They are also associated with a tendency to assume and maintain fixed postures. All aspects of movement are affected, including initiation, alteration in direction, and the ability to stop a movement once it is begun. Spontaneous or associated movements, such as swinging of the arms in gait or smiling at a funny story, are also affected. Bradykinesia is hypothesized to be the result of a decrease in activation of the supplementary motor cortex, premotor cortex, and motor cortex.104 The resting level of activity in these areas of the cortex may be decreased so that a greater amount of excitatory input from other areas of the brain would be necessary before movement patterns could be activated. In the individual with Parkinson disease, an increase in cortically initiated movement even for such “subcortical” activities as walking supports this hypothesis. Automatic activities are cortically controlled, and each individual aspect seems to be separately programmed. Associated movements in the trunk and other extremities are not automatic. This means that great energy must be expended whenever movement is begun.105 Bradykinesia and akinesia affect performance of all types of movements; however, complex movements are more involved than simple movements, such as dorsiflexing the foot at toe-off in walking as opposed to dorsiflexing the foot in a seated position.71,106–109 In addition, patients with parkinsonism have increased difficulty performing simultaneous or sequential tasks, over and above that seen with simple tasks. Parkinsonian patients must complete one movement before they can begin to perform the next, whereas control subjects are able to integrate two movements more smoothly in sequence. This deficit has been shown in a variety of tasks from performing an elbow movement and grip to tracing a moving line on a video screen. The patient with Parkinson disease behaves as if one motor program must be completely played out before the next one begins, and there is no advance planning for the next movement while the current movement is in progress.106–108,110,111 Morris and colleagues demonstrated a similar phenomenon in walking. Patients with parkinsonism were unable to perform walking while carrying a tray with a glass of water and had even more difficulty when walking and reciting a numerical sequence.112,113 Sequential movements become more impaired as more movements are strung together; for example, a square is disproportionately slower to draw than a triangle; a pentagon, more difficult than a square.5,106 These results indicate that patients with Parkinson disease have difficulty with transitions between movements. Transitional difficulties are more impaired in tasks requiring a series of different movements than tasks requiring a series of repetitive movements. For example, an individual will have less difficulty continually riding a stationary bike than movement requiring transitions such as coming from a chair to standing, walking, and turning a corner. Therefore treatment must include complex movements with directional changes to ensure that the patient is safe outside the treatment setting. Bradykinesia is not caused by rigidity or an inability to relax. This was demonstrated in an electromyographic analysis of voluntary movements of persons with Parkinson disease.114 Although the pattern of electromyographic agonist-antagonists burst is correct, these bursts are not large enough, resulting in an inability to generate muscle force rapidly enough. Even in slow, smooth movements, however, these individuals demonstrated alternating bursts in the flexor and extensor muscle groups. This type of pattern, expected in rapid movements that require the immediate activation of the antagonist to halt the motion, interferes with slow, smooth, continuous motion. Other researchers have found an alteration in the recruitment order of single motor units.115,116 These alterations included a delay in recruitment, pauses in the motor unit once it was recruited, and an inability to increase firing rates. These persons therefore would have a delay in activation of muscles and an inability to properly sustain muscle contraction for movement, and a decreased ability to dissipate force rapidly.24,115,117 Such changes may account for perceived decreases in strength that are seen in persons with Parkinson disease. They are also important to remember in both treatment planning and the efficacy of treatment efficiency. The rigidity (increased resistance to passive movement) of Parkinson disease may be characterized as either “lead pipe” or “cogwheel.” The cogwheel type of rigidity is a combination of lead-pipe rigidity with tremor. In rigidity there is an increased resistance to movement throughout the entire range in both directions without the classic clasp-knife reflex so characteristic of spasticity. Procaine injections can decrease the rigidity without affecting the decrease of spontaneous movements, confirming that rigidity is not the same phenomenon as bradykinesia.118,119 Rigidity is not caused by an increase in gamma motor neuron activity, a decrease in recurrent inhibition, or a generalized excitability in the motor system.120 Long- and middle-latency reflexes are enhanced in parkinsonism, and the increase in long-latency reflexes approximates the observable increase in muscle tone. Short-latency reflexes (i.e., deep tendon reflexes), on the other hand, may be normal in persons with Parkinson disease. Tatton and others121 found differences in certain cortical long-loop reflexes in normal and drug-induced parkinsonian monkeys, which led them to speculate that the “reflex gain” of the CNS may lose its ability to adjust to changing environmental situations. For example, in normal persons the background level of motor neuron excitability is different for the task of writing than for the task of lifting a heavy object; in individuals with Parkinson disease motor neuron excitability would be set at the same level. Similarly, in the normal individual there would be a difference in excitability if the environmental demands were for excitation or inhibition of a muscle; for the individual with Parkinson disease, there would be similar motor neuron excitability regardless of task demands. Furthermore, this lack of modulation may mean that the person with parkinsonism perceives himself or herself to be moving farther than he or she is actually moving. It is also consistent with a decrease in system flexibility and an inability to adjust to equilibrium perturbations.58,59,65 An important aspect of rigidity is that it might increase energy expenditure.122 This would increase the patient’s perception of effort on movement and may be related to feelings of fatigue, especially postexercise fatigue.123 The tremor observed in Parkinson disease is present at rest, usually disappears or decreases with movement, and has a regular rhythm of about 4 to 7 beats per second. Some people with Parkinson disease may have a postural tremor. The electromyographic tracing of a person with such a tremor shows rhythmical, alternating bursting of antagonistic muscles. Tremor can be produced as an isolated finding in experimental animals that have lesions in various parts of the brain stem or that have been treated with drugs, especially DA antagonists. DA depletion, however, is not the sole cause of tremor. It appears that efferent pathways, especially from the basal ganglia to the thalamus, must be intact because lesions of these fibers decrease or abolish the tremor.124 Poirier and colleagues124 proposed that tremor results from a combined lesion of the basal ganglia and cerebellar–red nucleus pathways. Because both the basal ganglia and the cerebellum project to the thalamus, a lesion of the thalamus can abolish the tremor regardless of the specific pathway(s). Although tremor may be cosmetically disabling, the tremor rarely interferes with activities of daily living (ADLs). Postural instability is a serious problem in parkinsonism that leads to increased episodes of falling and the sequelae of falls. More than two thirds of all patients with parkinsonism fall, and more than 10% fall more than once a week.125 People with Parkinson disease have a ninefold risk of recurrent falls compared with age-matched control subjects.60,126–130 Patients have an increased likelihood of falling as the duration of the disease increases. Drug treatment is not usually effective in reducing the incidence of falls. Deep brain stimulation and exercise, on the other hand, have been shown to be effective in increasing functional skills and/or motor performance; these improvements may decrease the number of falls.131–134 Large randomized clinical trials have been performed to determine the efficacy of exercise.135 Although the causes of balance difficulties are not known, several hypotheses exist. One explanation for postural instability is ineffective sensory processing. Several investigators have found deficits in proprioceptive and kinesthetic processing.55,74,117,136 For example, Martin55 found that labyrinthine equilibrium reactions were delayed in patients with Parkinson disease. Studies of the vestibular system itself, however, have shown that this system functions normally. Pastor and colleagues137 studied central vestibular processing in patients with Parkinson disease and found that the vestibular system responds normally and that patients can integrate vestibular input with the input from other sensory systems. This group hypothesized that the parkinsonian patients had an inability to adequately compensate for baseline instability. This theory is in partial agreement with studies by Beckley, Boehm, and others58,59,65 demonstrating that patients with Parkinson disease were unable to adjust the size of long- and middle-latency reflex responses to the degree of perturbation. These patients are therefore unable to activate muscle force proportional to displacement. Melnick and colleagues56 found that subjects with Parkinson disease were unable to maintain balance on a sway-referenced force plate. Glatt138 found that patients with Parkinson disease did not demonstrate anticipatory postural reactions and, in fact, behaved exactly as a rigid body with joints. Horak and colleagues,139,140 in a variety of studies, reported similar findings and found defects in strategy selection as well; patients with Parkinson disease chose neither a pure hip strategy nor a pure ankle strategy but mixed the two in an inappropriate and maladaptive response. Investigators have found that antiparkinsonian medications could improve background postural tone but did not improve automatic postural responses to external displacements.58,59,65,139–141 Other studies have demonstrated deficits in proprioceptive perception—what has been termed an “impaired proprioceptive body map.” Patients with Parkinson disease did not alter anticipatory postural adjustments in response to step width changes, unlike control subjects.142 Increased step width requires increased lateral reactive forces to unload the stance leg. The lack of ability to prepare for these extra forces may indicate that narrow stance width, start hesitation, and freezing of gait are compensatory mechanisms to proprioceptive loss.136 Likewise, when patients could not see their limbs, they had difficulty moving the foot to a predetermined location in response to perturbation. Control subjects had no difficulty.143,144 Taken together, it appears that postural instability results from inflexibility in response repertoire; an inability to inhibit unwanted programs; the interaction of akinesia, bradykinesia, and rigidity; and some disturbance in central sensory processing. The typical parkinsonian gait is characterized by decreased velocity and stride length.145,146 As a consequence, foot clearance is decreased, which again places the individual at greater fall risk.147 In many patients, especially as the disease progresses, speed and shortening of stride progressively worsen as if the individual is trying to catch up with his or her center of gravity; this is termed festination. Forward festination is called propulsion; backward festination is known as retropulsion. One hypothesis is that festinating gait is caused by the decreased equilibrium responses. If walking is a series of controlled falls and if normal responses to falling are delayed or not strong enough, then the individual will either fall completely or continue to take short, running-like steps. The abnormal motor unit firing seen with bradykinesia may also be the cause of ever-shortening steps. If the motor unit cannot build up a high enough frequency or if it pauses in the middle of the movement, then the full range of the movement would decrease; in walking this would lead to shorter steps. Festination may also be the result of other changes in the kinematics of gait. The changes in gait kinematics include changes in excursion of the hip and ankle joints (Figure 20-7). Instead of a heel-toe, the patient may have a flat-footed or, with disease progression, a toe-heel sequence. The patient with Parkinson disease appears to have lost the adult gait pattern and is using a more primitive pattern. The flat-footed gait decreases the ability to step over obstacles or to walk on carpeted surfaces. The use of three-dimensional gait analysis has shown that there is a decrease in plantarflexion at terminal stance. Changes are also seen in hip flexion, which may alter ankle excursion. However, qualitative aspects of the timing of joint excursion appear intact. Figure 20-7 illustrates the joint angles in a 55-year-old patient with Parkinson disease compared with adults without basal ganglia dysfunction.148 Especially in recent years, researchers have tried to address the cognitive and perceptual impairments of people with Parkinson disease.136,149–152 Whereas the movement deficits are hypothesized to be caused by a decrease in putaminal excitation of the cortex, the learning and perceptual deficits are hypothesized to be caused by a decrease in cortical excitation from the caudate nucleus.111 The deficits are of frontal lobe function and include an inability to shift attention, an inability to quickly access “working memory,” and difficulty with visuospatial perception and discrimination. Research attention has focused on the specific deficits of parkinsonian patients compared with patients with Alzheimer disease, patients with frontal lobe damage, and those with temporal lobe damage.149,152,153 The perceptual deficits of all groups appear to increase with progression of the disease process. In general, patients have difficulty in shifting attention to a previously irrelevant stimulus,154 learning under conditions requiring selective attention,154 or selecting the correct motor response on the basis of sensory stimuli.155–157 There is also evidence that DA is involved in selection of responses that will be rewarding.54 These impairments will affect treatment strategies. Learning deficits also have been found in patients with parkinsonism; procedural learning has been particularly implicated, as would be indicated based on the physiology of the system. Procedural learning is learning that occurs with practice or, as defined by Saint-Cyr and colleagues,158 “the ability gradually to acquire a motor skill or even a cognitive routine through repeated exposure to a specific activity constrained by invariant rules.” In their tests, patients with Parkinson disease did very poorly on tests of procedural learning, but their declarative learning was within normal limits. Pascual-Leone and colleagues111 studied procedural learning in more detail. They found that patients with Parkinson disease could acquire procedural learning but needed more practice than control subjects did. They also found that the ability to translate procedural knowledge to declarative knowledge was more efficient if it occurred with visual input alone rather than the combination of visual input with motor task. This may be a rationale for more therapy, not less.
Basal ganglia disorders
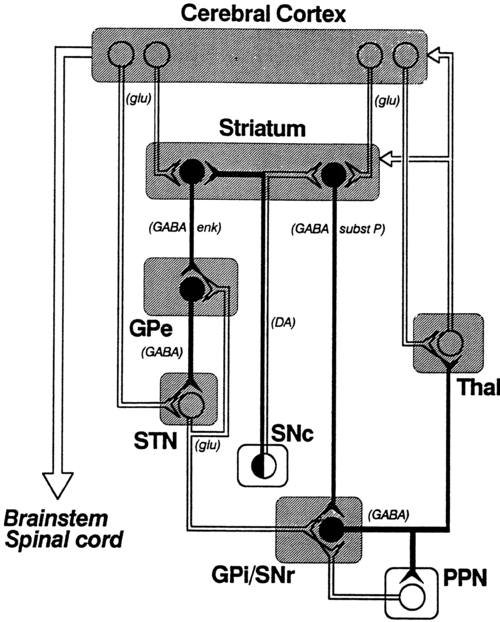
The basal ganglia
Anatomy
Afferent pathways
Efferent pathways
Pathways to the motor system
Physiology
Relationship of the basal ganglia to movement and posture
Automatic movement
Motor problems in animals
Movement initiation and preparation
Postural adjustments
Perceptual and cognitive functions
Neurotransmitters
Specific clinical problems arising from basal ganglia dysfunction
Parkinson disease
Symptoms
Bradykinesia and akinesia.
Rigidity.
Tremor.
Postural instability.
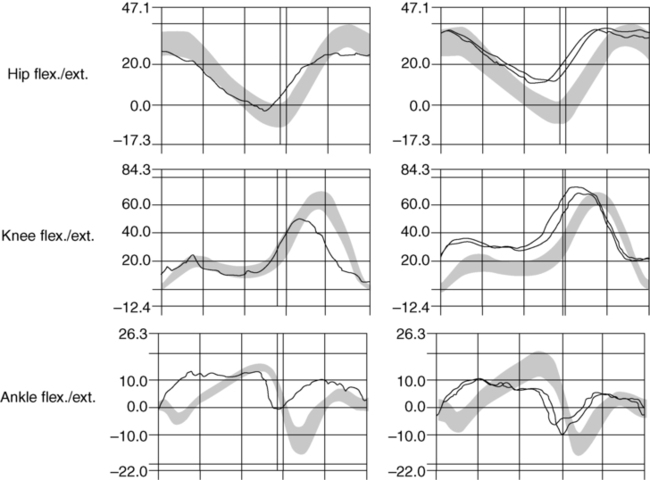
Gait.
Perception, attention, and cognitive deficits.
Related posts:
![]() Multiple sclerosis
Multiple sclerosis
![]() Contemporary issues and theories of motor control, motor learning, and neuroplasticity
Contemporary issues and theories of motor control, motor learning, and neuroplasticity
![]() The limbic system: influence over motor control and learning
The limbic system: influence over motor control and learning
![]() Movement analysis across the life span
Movement analysis across the life span
![]() Movement dysfunction associated with cerebellar damage
Movement dysfunction associated with cerebellar damage
![]() Neonates and parents: neurodevelopmental perspectives in the neonatal intensive care unit and follow-up
Neonates and parents: neurodevelopmental perspectives in the neonatal intensive care unit and follow-up
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Basal ganglia disorders









Only gold members can continue reading. Log In or Register to continue
