Chapter 1 Molecular Mechanisms of Cardiac Electrical Activity
Ionic Equilibrium
The lipid bilayer of the cell membrane is hydrophobic and impermeable to water-soluble substances such as ions. Hence, for the hydrophilic ions to be able to cross the membrane, they need hydrophilic paths that span the membrane (i.e., pores), which are provided by transmembrane proteins called ion channels. Once a hydrophilic pore is available, ions move passively across the membrane driven by two forces: the electrical gradient (voltage difference) and the chemical gradient (concentration difference). The chemical gradient forces the ions to move from a compartment of a higher concentration to one of lower concentration. The electrical gradient forces ions to move in the direction of their inverse sign (i.e., cations [positively charged ions] move toward a negatively charged compartment, whereas anions [negatively charged ions] move toward a positively charged compartment). Because the chemical and electrical gradients can oppose each other, the direction of net ion movement will depend on the relative contributions of chemical gradient and electrical potential (i.e., the net electrochemical gradient), so that ions tend to move spontaneously from a higher to a lower electrochemical potential.1–3
The movement of an ion down its chemical gradient in one direction across the cell membrane results in build-up of excess charge carried by the ion on one side of the membrane, which generates an electrical gradient that impedes continuing ionic movement in the same direction. When the driving force of the electrical gradient across the membrane becomes equal and opposite to the force generated by the chemical gradient, the ion is said to be in electrochemical equilibrium, and the net transmembrane flux (or current) of that particular ion is zero. In this setting, the electrical potential is called the equilibrium potential (Eion) (reversal potential or Nernst potential) of that individual ion. The Eion for a given ion depends on its concentration on either side of the membrane and the temperature, and it measures the voltage that the ion concentration gradient generates when it acts as a battery. At membrane voltages more positive to the reversal potential of the ion, passive ion movement is outward, whereas it is inward at a membrane potential (also known as transmembrane potential; Em) more negative to the Nernst potential of that channel.1,3
When multiple ions across a membrane are removed from their electrochemical equilibrium, each ion will tend to force the Em toward its own Eion. The contribution of each ion type to the overall Em at any given moment is determined by the instantaneous permeability of the plasma membrane to that ion. The larger the membrane conductance to a particular ion, the greater is the ability of that ion to bring the Em toward its own Eion. Hence, the Em is the average of the Eion of all the ions to which the membrane is permeable, weighed according to the membrane conductance of each individual ion relative to the total ionic conductance of the membrane.1,2
Transmembrane Potentials
All living cells, including cardiomyocytes, maintain a difference in the concentration of ions across their membranes. There is a slight excess of positive ions on the outside of the membrane and a slight excess of negative ions on the inside of the membrane, resulting in a difference in the electrical charge (i.e., voltage difference) across the cell membrane, called the membrane potential (Em). A membrane that exhibits an Em is said to be polarized.2
In nonexcitable cells, and in excitable cells in their baseline states (i.e., not conducting electrical signals), the Em is held at a relatively stable value, called the resting potential. All cells have a negative resting Em (i.e., the cytoplasm is electrically negative relative to the extracellular fluid), which arises from the interaction of ion channels and ion pumps embedded in the membrane that maintain different ion concentrations on the intracellular and extracellular sides of the membrane.2
When an ion channel opens, it allows ion flux across the membrane that generates an electrical current (I). This current affects the Em, depending on the membrane resistance (R), which refers to the ratio between the Em and electrical current, as shown in Ohm’s law: E = I × R or R = E/I. Resistance arises from the fact that the membrane impedes the movement of charges across it; hence, the cell membrane functions as a resistor. Conductance describes the ability of a membrane to allow the flux of charged ions in one direction across the membrane. The more permeable the membrane is to a particular ion, the greater is the conductance of the membrane to that ion. Membrane conductance (g) is the reciprocal of R: g = 1/R.1
Because the lipid bilayer of the cell membrane is very thin, accumulation of charged ions on one side gives rise to an electrical force (potential) that pulls oppositely charged particles toward the other side. Hence, the cell membrane functions as a capacitor. Although the absolute potential differences across the cell membrane are small, they give rise to enormous electrical potential gradients because they occur across a very thin surface. As a consequence, apparently small changes in Em can produce large changes in potential gradient and powerful forces that are able to induce molecular rearrangement in membrane proteins, such as those required for opening and closing ion channels embedded in the cell membrane. The capacitance of the membrane is generally fixed and unaffected by the molecules that are embedded in it. In contrast, membrane resistance is highly variable and depends on the conductance of ion channels embedded in the membrane.2,3
The sodium (Na+), potassium (K+), calcium (Ca2+), and chloride (Cl−) ions are the major charge carriers, and their movement across the cell membrane creates a flow of current that generates excitation and signals in cardiac myocytes. The electrical current generated by the flux of an ion across the membrane is determined by the membrane conductance to that ion (gion) and the potential (voltage) difference across the membrane. The potential difference represents the potential at which there is no net ion flux (i.e., the Eion) and the actual Em: current = gion × (Em − Eion).1,4
By convention, an inward current increases the electropositivity within the cell (i.e., causes depolarization of the Em [to be less negative]) and can result from either the movement of positively charged ions (most commonly Na+ or Ca2+) into the cell or the efflux of negatively charged ions (e.g., Cl−) out of the cell. An outward current increases the electronegativity within the cell (i.e., causes hyperpolarization of the Em [to become more negative]) and can result from either the movement of anions into the cell or the efflux of cations (most commonly K+) out of the cell.3
Opening and closing of ion channels can induce a departure from the relatively static resting Em, called a depolarization if the interior voltage rises (becomes less negative) or a hyperpolarization if the interior voltage becomes more negative. The most important ion fluxes that depolarize or repolarize the membrane are passive (i.e., the ions move down their electrochemical gradient without requiring the expenditure of energy), occurring through transmembrane ion channels. In excitable cells, a sufficiently large depolarization can evoke a short-lasting all-or-none event called an action potential, in which the Em very rapidly undergoes specific and large dynamic voltage changes.1
Both resting Em and dynamic voltage changes such as the action potential are caused by specific changes in membrane permeabilities for Na+, K+, Ca2+, and Cl−, which, in turn, result from concerted changes in functional activity of various ion channels, ion transporters, and exchangers.3
The Cardiac Action Potential
During physiological electrical activity, the Em is a continuous function of time. The current flowing through the cell membrane is, at each instant, provided by multiple channels and transporters carrying charge in opposite directions because of their different ion selectivity. The algebraic summation of these contributions is referred to as net transmembrane current.1
The cardiac action potential reflects a balance between inward and outward currents. When a depolarizing stimulus (typically from an electric current from an adjacent cell) abruptly changes the Em of a resting cardiomyocyte to a critical value (the threshold level), the properties of the cell membrane and ion conductances change dramatically, precipitating a sequence of events involving the influx and efflux of multiple ions that together produce the action potential of the cell. In this fashion, an electrical stimulus is conducted from one cell to all the cells that are adjacent to it.2
Unlike skeletal muscle, cardiac muscle is electrically coupled so that the wave of depolarization propagates from one cell to the next, independent of neuronal input. The heart is activated by capacitive currents generated when a wave of depolarization approaches a region of the heart that is at its resting potential. Unlike ionic currents, which are generated by the flux of charged ions across the cell membrane, capacitive currents are generated by the movement of electrons toward and away from the surfaces of the membrane.2,3 The resulting decrease in positive charge at the outer side of the cell membrane reduces the negative charge on the intracellular surface of the membrane. These charge movements, which are carried by electrons, generate a capacitive current. When an excitatory stimulus causes the Em to become less negative and beyond a threshold level (approximately −65 mV for working atrial and ventricular cardiomyocytes), Na+ channels activate (open) and permit an inward Na+ current (INa), resulting in a rapid shift of the Em to a positive voltage range. This event triggers a series of successive opening and closure of selectively permeable ion channels. The direction and magnitude of passive ion movement (and the resulting current) at any given transmembrane voltage are determined by the ratio of the intracellular and extracellular concentrations and the reversal potential of that ion, with the net flux being larger when ions move from the more concentrated side.3
The threshold is the lowest Em at which opening of enough Na+ channels (or Ca2+ channels in the setting of nodal cells) is able to initiate the sequence of channel openings needed to generate a propagated action potential. Small (subthreshold) depolarizing stimuli depolarize the membrane in proportion to the strength of the stimulus and cause only local responses because they do not open enough Na+ channels to generate depolarizing currents large enough to activate nearby resting cells (i.e., insufficient to initiate a regenerative action potential). On the other hand, when the stimulus is sufficiently intense to reduce the Em to a threshold value, regenerative action potential results, whereby intracellular movement of Na+ depolarizes the membrane more, a process that increases conductance to Na+ more, which allows more Na+ to enter, and so on. In this fashion, the extent of subsequent depolarization becomes independent of the initial depolarizing stimulus, and more intense stimuli do not produce larger action potential responses; rather, an all-or-none response results.2
During the action potential, membrane voltages fluctuate in the range of −94 to +30 mV (Fig. 1-1). With physiological external K+, the reversal potential of K+ (EK) is approximately −94 mV, and passive K+ movement during an action potential is out of the cell. On the other hand, because the calculated reversal potential of a cardiac Ca2+ channel (ECa) is +64 mV, passive Ca2+ flux is into the cell.5
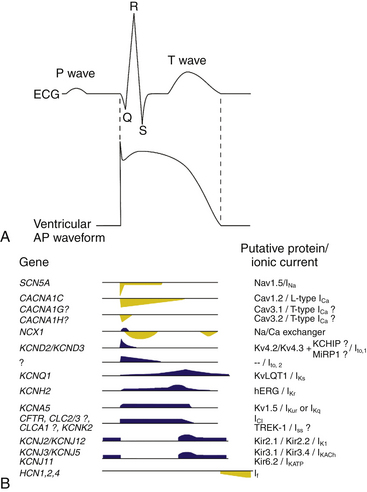
In normal atrial and ventricular myocytes and in His-Purkinje fibers, action potentials have very rapid upstrokes, mediated by the fast inward INa. These potentials are called fast response potentials. In contrast, action potentials in the normal sinus and atrioventricular (AV) nodal cells and many types of diseased tissues have very slow upstrokes, mediated by a slow inward, predominantly L-type voltage-gated Ca2+ current (ICaL), rather than by the fast inward INa (Fig. 1-2). These potentials have been termed slow response potentials.2,5
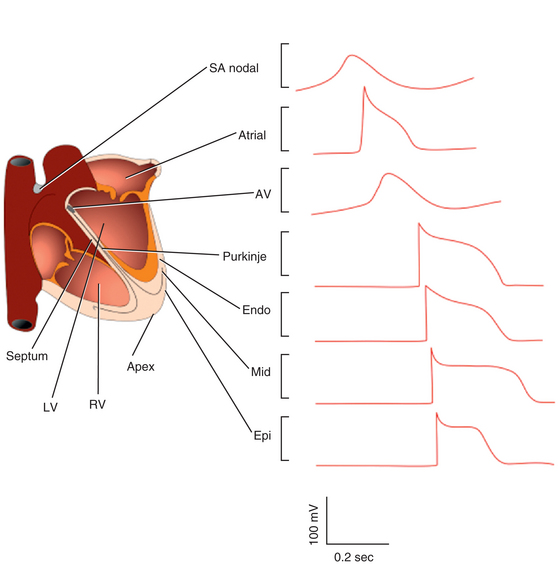
The Fast Response Action Potential
Phase 4: The Resting Membrane Potential
The Em of resting atrial and ventricular cardiomyocytes remains steady throughout diastole. The resting Em is caused by the differences in ionic concentrations across the membrane and the selective membrane permeability (conductance) to various ions. Large concentration gradients of Na+, K+, Ca2+, and Cl− across the cell membrane are maintained by the ion pumps and exchangers (Table 1-1).
TABLE 1-1 Intracellular and Extracellular Ion Concentrations and Equilibrium Potentials in Cardiomyocytes
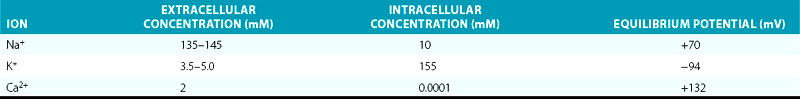
Under normal conditions, the resting membrane is most permeable to K+ and relatively impermeable to other ions. K+ has the largest resting membrane conductance (gK is 100 times greater than gNa) because of the abundance of open K+ channels at rest, whereas Na+ and Ca2+ channels are generally closed. Thus, K+ exerts the largest influence on the resting Em. As a consequence, the resulting Em is almost always close to the K+ reversal potential (Em approximates EK). The actual resting Em is slightly less negative than EK because the cell membrane is slightly permeable to other ions.2
The inwardly rectifying K+ (Kir) channels underlie an outward K+ current (IK1) responsible for maintaining the resting potential near the EK in atrial, His-Purkinje, and ventricular cells, under normal conditions.6,7 Kir channels preferentially allow currents of K+ ions to flow into the cell with a strongly voltage-dependent decline of K+ efflux (i.e., reduction of outward current) on membrane depolarization. As such, IK1 is a strong rectifier that passes K+ currents over a limited range of Em (see Chap. 2 for detailed discussion on the concept of rectification); at a negative Em, IK1 conductance is much larger than that of any other current, thus, it clamps the resting Em close to the reversal potential for K+ (EK). IK1 density is much higher in ventricular than in atrial myocytes, a feature that protects the ventricular cell from pacemaker activity. By contrast, IK1 is almost absent in sinus and AV nodal cells, thus allowing for relatively more depolarized resting diastolic potentials compared with atrial and ventricular myocytes (Table 1-2).2

A unique property of Kir currents is the unusual dependence of rectification on extracellular K+ concentration. Specifically, on increase in extracellular K+, the IK1 current-voltage relationship shifts nearly in parallel with the EK and leads to a crossover phenomenon. One important consequence of such behavior is that at potentials positive to the crossover, K+ conductance increases rather than decreases, against an expectation based on a reduced driving force for K+ ions as a result of elevated extracellular K+ concentration.6–8
Although Ca2+ does not contribute directly to the resting Em (since the voltage-activated Ca2+ channels are closed at the hyperpolarized resting Em), changes in intracellular free Ca2+ concentration can affect other membrane conductance values. Increases in intracellular Ca2+ levels can stimulate the Na+-Ca2+ exchanger (INa-Ca), which exchanges three Na+ ions for one Ca2+ ion; the direction depends on the Na+ and Ca2+ concentrations on the two sides of the membrane and the Em difference. At resting Em and during a spontaneous sarcoplasmic reticulum Ca2+ release event, this exchanger would generate a net Na+ influx, possibly causing transient membrane depolarizations.3
Phase 0: The Upstroke—Rapid Depolarization
On excitation of the cardiomyocyte by electrical stimuli from adjacent cells, its resting Em (approximately −85 mV) depolarizes, leading to opening (activation) of Na+ channels from its resting (closed) state and enabling a large and rapid influx of Na+ ions (inward INa) into the cell down their electrochemical gradient. As a consequence of increased Na+ conductance, the excited membrane no longer behaves like a K+ electrode (i.e., exclusively permeable to K+), but more closely approximates an Na+ electrode, and the membrane potential moves toward the Na+ Eion (ENa; see Table 1-1). Once an excitatory stimulus depolarizes the Em beyond the threshold for activation of Na+ channels (approximately −65 mV), the activated INa is regenerative and no longer depends on the initial depolarizing stimulus; the influx of Na+ ions further depolarizes the membrane and thereby increases conductance to Na+ more, which allows more Na+ to enter.2,9
Normally, activation of Na+ channels is transient; fast inactivation (closing of the pore) starts simultaneously with activation, but because inactivation is slightly delayed relative to activation, the channels remain transiently (less than 1 millisecond) open to conduct INa during phase 0 of the action potential before it closes. Additionally, the influx of Na+ into the cell increases the positive intracellular charges and reduces the driving force for Na+. When the ENa is reached, no further Na+ ions enter the cell.9
The rate at which depolarization occurs during phase 0, that is, the maximum rate of change of voltage over time, is indicated by the expression dV/dtmax, which is a reasonable approximation of the rate and magnitude of Na+ entry into the cell and a determinant of conduction velocity for the propagated action potential.2
The threshold for activation of ICaL is approximately −30 to −40 mV. Although ICaL is normally activated during phase 0 by the regenerative depolarization caused by the fast INa, ICaL is much smaller than the peak INa. The amplitude of ICaL is not maximal near the action potential peak because of the time-dependent nature of ICaL activation, as well as the low driving force (Em − ECa) for ICaL. Therefore, ICaL contributes little to the action potential until the fast INa is inactivated, after completion of phase 0. As a result, ICaL affects mainly the plateau of action potentials recorded in atrial and ventricular muscle and His-Purkinje fibers. On the other hand, ICaL can play a prominent role in the upstroke of slow response action potentials in partially depolarized cells in which the fast Na+ channels have been inactivated.2






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


