4
General Anatomy, Physiology, and Biomechanics
| Development of the Intervertebral Disk | 15 | Biomechanics | 22 |
| Anatomy | 18 | The Disk and the Intervertebral Joints | 29 |
| Fine Structure and Biochemistry | 21 | Therapeutic Approaches | 31 |
 Development of the Intervertebral Disk
Development of the Intervertebral Disk
An understanding the pathoanatomical changes affecting the intervertebral disk in adulthood requires knowledge of its developmental history in utero and in early childhood. Even at these very early stages of development, evidence can often be found indicating that the process of disk degeneration—which affects humans at a relatively young age—has already begun. The more important developmental studies of the normal and abnormal spine are those of Töndury (1947, 1955, 1958, 1968, 1970, 1981) and his colleagues Larcher (1947), Prader (1947), and Ecklin (1960), as well as those of Bell et al. (1990) and Bogduk (2000). These researchers were able to demonstrate, in serial histological sections, the regression of the notochord and the development of the vertebral bodies and primordial intervertebral disks (Figs. 4.1–4.3).
An axial cord of cells that is at first only one cell thick, called the notochord (from the Greek notos, “back”) or chorda dorsalis, is found in the embryos of all chordate animals and gives them their name. In lancelets (Cephalochordata) and tunicates (Urochordata, sea squirts), the notochord remains the only axial skeleton for the entire life of the organism. In vertebrates, the notochord disappears at an early stage of embryonic development and is replaced by the cartilaginous or bony spine. Only a few weeks after conception, the embryo (measuring 12 mm from vertex to sacrum) contains a spinal column with distinct vertebrae and intervertebral disks; these are pierced by the notochord, which is still present along the entire length of the spine (Fig. 4.1).
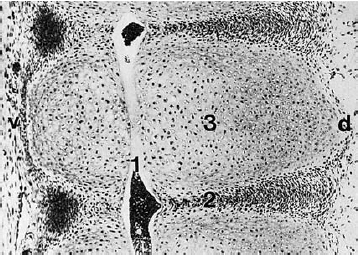
Fig. 4.1 Midline sagittal section of the primitive vertebral column of a 12 mm embryo. The notochord (1) runs craniocaudally through the primordial intervertebral disks (2) and vertebral bodies (3). v = ventral, d = dorsal. (Original photograph courtesy of Prof. Christ, Anatomical Institute, Univ. of Bochum, Germany.).
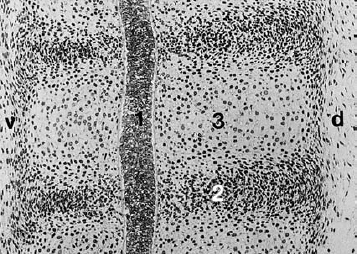
Fig. 4.2 In the 30 mm embryo, the notochord (1) is thickened at the level of the intervertebral disk (2). During the course of further development, the notochord normally disappears entirely from the primordial vertebral bodies (3). d = dorsal. (Original photograph courtesy of Prof. Christ, Anatomical Institute, Univ. of Bochum, Germany.).
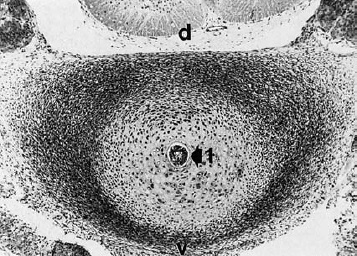
Fig. 4.3 Transverse section of the disk in a 30 mm embryo. The notochord (1) lies at the center. The peripheral, thickened portion of the disk blastema will develop into the anulus fibrosus. v = ventral, d = dorsal. (Original photograph courtesy of Prof. Christ, Anatomical Institute, Univ. of Bochum, Germany.).
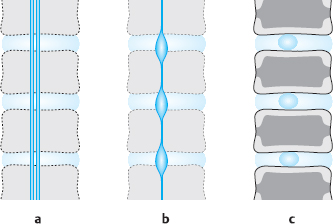
Fig. 4.4a–c Development of the intervertebral disk.
a 12 mm embryo. Primordial vertebral column with cartilaginous vertebral bodies and intervertebral disks with longitudinally running fibrils in the outer zone; the notochord traverses all segments.
b 50mm embryo. The notochord has been pressed out of the vertebral bodies, and remnants of it are found only in the intervertebral disks.
c 6-year-old child. Notochord remnants are found only in the central portion of the intervertebral disks. Central ossification of the vertebral bodies. The transitional zone between the vertebral body and the intervertebral disk forms plates of cartilage with cartilaginous margins that later give rise to bony margins (cf.Fig. 4.5 a–c).
Later in development, the notochord is squeezed out of the primordial vertebral bodies by the growing cartilage cells, so that it ultimately persists only as droplike remnants in the center of the intervertebral disks, marking the location of what will later be the nucleus pulposus. The primordial disk tissue surrounding the notochordal remnant consists of outer and inner zones. The outer zone will later become the anulus fibrosus. Even at an early stage of development, it already contains longitudinally running fibers that radiate into the cartilaginous layer of the primordial vertebral body. These are the forerunners of Sharpey’s fibers in the transitional zone between disk and vertebra. The outer zone contains many fibers and few cells; it undergoes a smooth transition to the pulpy inner zone around the notochord, which contains fewer structures (Fig. 4.4).
The parachordal inner zone and the somewhat eccentrically situated notochordal remnant together give rise to the nucleus pulposus. While the center of the vertebral body gradually ossifies, a cartilaginous plate forms at the interface of the vertebral body and the disk. The cartilaginous margin of this plate will later give rise to the bony margin of the vertebral body.
All of the disk structures destined to play a role in the biomechanical function of the spine are already present at birth.
 The growing intervertebral disk still possesses its own blood supply during embryonic development and infancy.
The growing intervertebral disk still possesses its own blood supply during embryonic development and infancy.
These blood vessels arise in the vascular network lying just outside the spine, particularly in the intervertebral foramina, and travel directly into the anulus fibrosus, radially penetrating its layers (lamellae) and forming interlamellar capillary networks. There are two types of blood vessels that supply the disk: peripheral and central axial. They never penetrate into the inner layers of the anulus fibrosus or into the nucleus pulposus. Thus, from the very beginning of its development the central portion of the intervertebral disk is nourished exclusively by diffusion.
The vertebral bodies and the intervertebral disks do not reach their final form till early adulthood. The vertebral bodies grow from the proliferative zones of the cartilaginous end plates. The side of the end plate facing the bone marrow bears a typical zone of cartilage growth and degradation that remains present till about age 20.
Areas of ossification appear within the ring of cartilage at the edge of the cartilaginous end plate, joining together at about age 12 to form a bony ring which then fuses with the vertebral body. The lamellae of the anulus fibrosus are anchored to this bony marginal ring by Sharpey’s fibers (Fig. 4.5).
The anulus fibrosus and the nucleus pulposus increase in size by interstitial appositional growth. Dense lamellar bundles form in the outer layers of the disk and pass between the two vertebral bodies in variably intertwined helical configurations.
The lamellae of the anulus fibrosus are weaker and less densely packed the further they are from the periphery of the disk; thus, even when the disk is fully mature, its center—the nucleus pulposus—consists mostly of structureless matrix. Töndury (1955) and Schaaf et al. (2004) found that the connective tissue cells in the anulus fibrosus, which produce fibers and matrix, receive a blood supply only until about the second year of life. The nourishing vessels regress thereafter, so that the anulus fibrosus is avascular even in a 4-year-old child. The function of this vascular regression is not known. On the contrary, the human intervertebral disk, rich as it is in cells and fibers, might seem to require an abundant blood supply, in view of the continual synthesis and degradation of macromolecules taking place within it. The regression of the blood supply of the intervertebral disk roughly coincides with the assumption of an upright posture at the age of about 1 year. One may suspect a connection between the avascularity of the disk and the mechanical stresses continually placed on it by standing and walking.
The arteries and veins of the vertebrae lie in the empty spaces left by the hard, bony trabecular system and are thus not exposed to axial mechanical stress. In contrast, the pulpy, homogeneous tissue of the interior of the disk physically resembles a fluid. Thus, depending on body position, the intradiscal pressure can be high enough to cause prolonged compression of the intradiscal vessels, leading in turn to an impairment of metabolism in the disk tissue.
 The reduced supply of nutrients to the intervertebral disk adversely affects both the quantity and the quality of the intradiscal connective and supporting tissues.
The reduced supply of nutrients to the intervertebral disk adversely affects both the quantity and the quality of the intradiscal connective and supporting tissues.
In the ensuing years, the anulus fibrosus and the nucleus pulposus increase in volume by interstitial appositional growth, but their growth is outstripped by that of the vertebral body. Thus, the ratio of vertebral height to disk height gradually increases from 1:1 at birth to 3:1–5:1 by the end of the axial growth phase. Qualitatively, too, the intervertebral disks of adolescents display involutional changes suggesting premature aging, mainly because of a rapid loss of water content.
Changes in the consistency and color of the disk tissue over the first few years of life can easily be seen with the naked eye in fresh autopsy specimens. In neonates and infants, the cut surface of the intervertebral disk appears glassy, gelatinous, and semi-liquid (Fig. 4.6). Even in a 2-year-old, for example, one can wipe away the semi-liquid central portion of the disk with a cotton swab, but in the adult disk this is no longer possible.
Even after the completion of axial growth, the disk undergoes further regressive changes that alter its appearance. In advanced age, the central disk tissue loses its original, homogeneous, gelatinous character and instead looks dry and fibrous. If the motion segment is immobilized by spondylotic spurs, the connective tissue of the disk may become reorganized, and there may be renewed ingrowth of blood vessels into the disk. Hassler (1969) regularly found granulation tissue and blood vessels inside severely degenerated disks.
 The intervertebral disk continues to change throughout life, from infancy to old age, and is particularly vulnerable to disease in certain phases of life. These are the times at which disk-related symptoms tend to arise.
The intervertebral disk continues to change throughout life, from infancy to old age, and is particularly vulnerable to disease in certain phases of life. These are the times at which disk-related symptoms tend to arise.
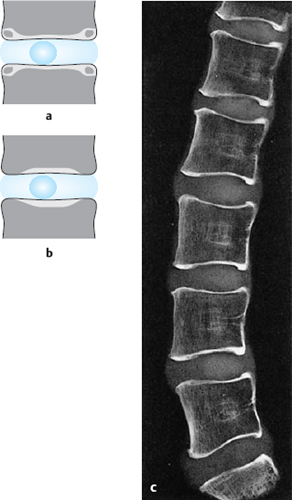
Fig. 4.5 a–c Development of the interface between the vertebral body and the intervertebral disk.
a 12-year-old child. A bony ring, the bony margin, forms within the cartilaginous margin between the ages of 8 and 12.
b Adult. The bony margin fuses with the vertebral body at the age of 14 or 15.
c Lateral radiograph of a midline sagittal section of a 17-year-old boy (saw-cut anatomical preparation).
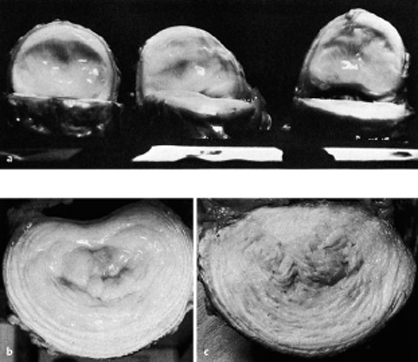
Fig. 4.6 a–c Lumbar intervertebral disks in cross-section.
a 2-year-old boy.
b 15-year-old boy.
c 42-year-old man.
 Anatomy
Anatomy
The conditions that are known as intervertebral disk disease involve pathological changes not only in the disks themselves, but also in the neighboring tissues, which have closely related functions. In this section, we discuss the normal anatomy in and around the intervertebral disks only in as much detail as necessary for an understanding of pathological changes in these areas. The particular anatomical features of the different regions of the spine will be described in the corresponding chapters.
 The motion segment is the building block and functional unit of the spine.
The motion segment is the building block and functional unit of the spine.
One important component of the motion segment is the intervertebral disk, comprising the nucleus pulposus, the anulus fibrosus, and the cartilaginous end plates (though the end plates belong, embryologically speaking, to the vertebral bodies). Its other components are the lower half of the upper vertebral body and the upper half of the lower vertebral body, the anterior and posterior longitudinal ligaments, the ligamentum flavum (yellow ligament), the intervertebral joints, and all of the other soft tissues found at the level of the motion segment, in the spinal canal, in the intervertebral foramen, and between the spinous and transverse processes.
The human spine generally comprises 25 motion segments, of which the upper two (atlanto-occipital and atlantoaxial) do not possess an intervertebral disk. The disks are named according to the neighboring vertebral bodies. There are normally 23 of them: 5 cervical, 11 thoracic, 4 lumbar, and one each at the junctions between the regions of the spine—cervicothoracic, thoracolumbar, and lumbosacral. The lowest disk (L5/S1) is also sometimes called the presacral disk. Individuals with 6 lumbar vertebrae will correspondingly have 5 lumbar disks (Figs. 4.7, 4.8). In the adult, the disks account for one-quarter of the length of the spine.
The intervertebral disks increase in height going down the spine. They have a trapezoidal shape in the sagittal plane, corresponding to the normal spinal curvatures. The normal convexity of the cervical and lumbar lordoses is due to the greater height of the anterior portion of the cervical and lumbar disks, but the normal thoracic kyphosis is due to the shapes of the vertebral bodies.
 The term “intervertebral disk” refers to all of the nonosseous tissues lying between two adjacent vertebral bodies: the cartilaginous end plates, the anulus fibrosus, and the nucleus pulposus.
The term “intervertebral disk” refers to all of the nonosseous tissues lying between two adjacent vertebral bodies: the cartilaginous end plates, the anulus fibrosus, and the nucleus pulposus.
The fibrous ring (anulus fibrosus) of the intervertebral disk consists mainly of fibers that are interwoven with each other as they take a helically winding path from one vertebral body to the next. Its peripheral zone contains the tough Sharpey’s fibers, which are tightly anchored to the bony margins of the vertebral body. The lamellae of the anulus fibrosus are tougher and more numerous ventrally and laterally than dorsally and dorsolaterally; at the latter sites, the anulus fibrosus is thin, composed of only a few, relatively thin lamellae. The transition between the anulus fibrosus and the nucleus pulposus is gradual, without any sharply defined border between the two structures.
The nucleus pulposus is built up around the notochordal remnant and therefore does not lie in the exact center of the intervertebral disk, but somewhat more posteriorly. While the spine is still growing longitudinally, the nucleus pulposus contains not just the bubblelike cells of the notochordal remnant, but also the socalled notochordal reticulum, composed of reticulated strands of nuclei that look like notochord cell nuclei, yet lie directly next to one another. The interstices of the reticulum are filled with a slippery fluid matrix. A widely branching cavity is thus generated, which is at first filled with fluid resembling synovial fluid, and later with pulpy tissue that wells up as soon as the disk is cut open (Figs. 4.9, 4.10).
In advanced age, the tissue of the nucleus pulposus is easier to push apart, and it contains cavities. It is thus easier to inject 1–2 mL of fluid into the disk of an elderly person than into that of a young adult, whose disk tissue is still highly coherent.
When the intervertebral disk of a young adult is incised, the pulpy tissue of the interior of the disk immediately wells up out of the plane of the cut surface. The reason for this is that, once the nucleus is freed of the constraining framework of the anulus fibrosus and the vertebral bodies above and below, its internal pressure, i.e., its tissue turgor, leads it to assume a more spherical shape.
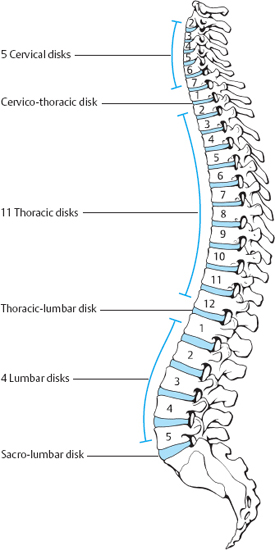
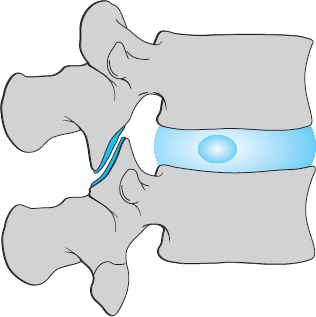
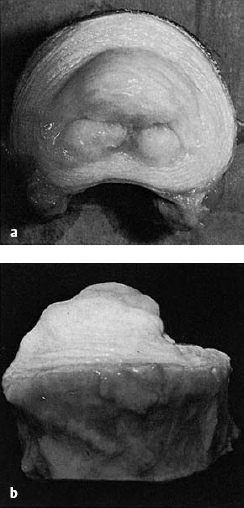
Fig. 4.9 a, b Lumbar disk of a 17-year-old boy. As soon as the disk is bisected, the nucleus pulposus wells up out of the plane of section.
Fig. 4.10 Lumbosacral disk of an 18-year-old man. The mobile tissue in the center of the disk is easily extracted with forceps.
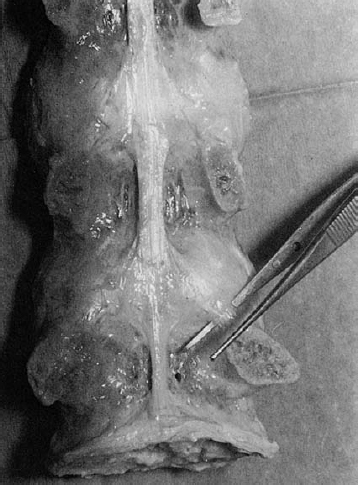
Fig. 4.11 Posterior longitudinal ligament in a dissected specimenofthelumbarspine.Theligamentwidensatthelevel of the disk, and some of its fibers course obliquely downward to the root of the lamina. The ligament does not cover the upper lateral portion of the posterior aspect of the disk.
Ligaments
The anterior longitudinal ligament is a broad ligamentous band that covers the anterior surfaces of the vertebral bodies and of the anuli fibrosi of the intervertebral disks. It can easily be dissected free of the underlying anular tissue.
The posterior longitudinal ligament, on the other hand, is tightly bound to the underlying anulus fibrosus and is more difficult to separate from it. It is broader cranially than caudally; at the lumbar vertebral level, it is no more than a narrow strip (Fig. 4.11). Our anatomical studies (Stahl 1977) have revealed that, despite what many anatomy texts say, the posterior longitudinal ligament does not entirely cover the posterior aspect of the lumbar disks. Their superolateral portion remains uncovered, and it is presumably no coincidence that disk prolapses are most common at this site. The lateral fibers of the posterior longitudinal ligament run obliquely downward at the level of the disk to terminate on the periosteum of the root of the lamina. Tension on these fibers, from a disk protrusion or other cause, can give rise to periosteal pain.
 Fine Structure and Biochemistry
Fine Structure and Biochemistry
Apart from the sparse remnants of the embryonic notochord, the human intervertebral disk contains only tissue components that are found in supportive and connective tissues throughout the body. The disk is clearly distinct from the structures around it, and its components are arranged in the intervertebral space in such a way that the mechanical properties of each are ideally suited to the stress occurring locally. Thus, the disk can be thought of as an organ composed of connective tissue. Its histologically and biochemically distinct components occupy different spatial positions to fulfill their respective biomechanical roles.
 The anulus fibrosus consists mainly of fibers, the nucleus pulposus of matrix, and the cartilaginous end plates of hyaline cartilage.
The anulus fibrosus consists mainly of fibers, the nucleus pulposus of matrix, and the cartilaginous end plates of hyaline cartilage.
These tissues are generated by connective tissue cells, which themselves make up between 20% and 30% of the overall tissue volume. The disk contains fibroblasts, cartilage cells, and a few notochord cells. These connective tissue cells produce matrix and intra- and extracellular fibers. To synthesize extracellular macromolecules, the cells need low-molecular-weight metabolic substrates, including amino acids, salts, glucose, and water. The cell density within the disk is a function of the local nutrient supply; the center of the disk thus contains fewer cells (Stairmand et al. 1991, Ito et al. 2001, Kluba et al. 2005).
 The water content of the nucleus pulposus decreases from 90% in the first year of life to 74% in the eighth decade.
The water content of the nucleus pulposus decreases from 90% in the first year of life to 74% in the eighth decade.
Our studies have shown that the water content varies in different parts of the anulus fibrosus (Kolditz et al. 1985, Krämer et al. 1985). The water is mainly present, not in free form, but rather as a structural component of macromolecules. It is reversibly bound to the free ionized groups on macromolecules; it can be exchanged for the hydrophilic groups of certain substances and thereby move into the interstitial fluid. The water content of the disks can be determined quantitatively by MRI (Panagiotacopoulos et al. 1987, Silcox 1995, Rajasekaran 2004). Our radiologists quantitatively measured the change in water content of the disks from morning to evening, as manifested by changing echo times (Beyer et al. 1986).
In addition to interstitial fluid, the disk tissue contains minerals, enzymes, organic matrix, and a small amount of fat. It does not normally contain a separate mineral phase; crystals of apatitic calcium phosphates are present only in elderly individuals. McCarty (1964) found brushite and calcium phosphate crystals in human disk tissue. Inorganic ions such as sodium, potassium, and calcium are partly structurally bound, partly dissolved in the extracellular fluid. Calcium ions are bound to acidic mucopolysaccharides in the matrix, where their concentration may be up to 35 times as high as in the extracellular fluid. The disk is thus rightly called a calcium-trapping organ. Mineralization occurs in tandem with a rise in phosphorus concentration and, often, a gradual precipitation of crystals. The high calcium content of the cartilaginous portion of the disk is accounted for by intracellular calcium. Sodium is partly bound to the matrix; all other ions are found in the interstitial fluid.
The organic matrix content of the disk increases from periphery to center, i.e., from the anulus fibrosus to the nucleus pulposus. The matrix contains mainly glycoproteins and high-molecular-weight polysaccharides. The glycoproteins consist of protein and carbohydrate; being mucoprotein secretions, they are viscous and highly hydrophilic. The high-molecular-weight polysaccharides of the matrix are mainly acidic mucopolysaccharides, such as hyaluronic acid, chondroitin sulfate, keratan sulfate, and heparin. These molecules form a highly polymerized, three-dimensional grid that lends viscosity to the matrix.
 By virtue of their high effective hydrodynamic volume, the macromolecules bind a large fraction of the fluid content of the intervertebral disk.
By virtue of their high effective hydrodynamic volume, the macromolecules bind a large fraction of the fluid content of the intervertebral disk.
Related posts:


Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


