Chapter 1 Cartilage Morphology
Hyaline cartilage provides the diarthrodial joint with a low-friction surface, resilience, and compressive stiffness, and this unique tissue is, under normal conditions, wear resistant.
Loss of cartilage function may lead to a painful joint with a decreased mobility. Many factors (epidemiological, biochemical, and morphological) are associated with cartilage destruction. However, only trauma is known directly to cause osteoarthritis.1 It is well known that once the cartilaginous tissue has been destroyed, the intrinsic reparative ability is poor. Therefore, it is of uttermost importance to increase knowledge about the cartilage, the tissue reaction to trauma, and the intrinsic attempts to repair the defects as well as extrinsic methods.
Cartilage Biochemistry and Morphology
The collagen fibers are responsible for the structure of cartilage and consist mainly of collagen type II. They are highly cross-linked via collagen type IX fibers.2
Chondrocytes are the producers of the surrounding ground substance: matrix.
The cells have different appearances depending on where in the cartilage they are situated. The cells in the top layer appear flattened, whereas the cells in the deeper layer are more rounded and aligned along vertically orientated type II collagen.3
Proteoglycans are large protein-polysaccharide molecules making up 5% to 10% of the wet weight of the cartilage.4 They are composed by chains of the glycosaminoglycans keratan sulfate and chondroitin sulfate covalently bound to a central protein core molecule. Large aggregates are formed with several proteoglycan monomers via a link protein connecting the central protein cores to a chain of hyaluronic acid.
All the components of the proteoglycan aggregates are synthesized by the chondrocytes.
The proteoglycans are unevenly distributed throughout the cartilage layers with the highest concentration in the middle part and the lowest concentration in the superficial layers.5 The proteoglycans give the cartilage its elasticity and resilience.
There is a difference in cartilage composition between the cartilage surface and the subchondral bone plate. These structural differences give rise to four separate layers or zones (see Fig. 1-1).
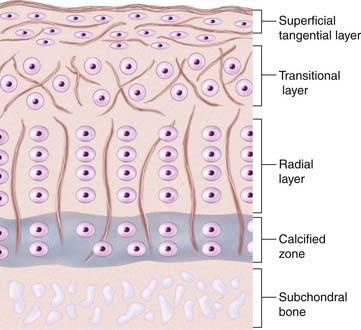
In the top zone, the superficial zone, there is first a cell-free fibril-layer, called the lamina splendens.6 Beneath this thin layer, chondrocytes are dispersed in an elongated manner parallel to the surface, reflecting as well the tangential orientation of the collagen fibers. This is the tangential layer.
The calcified zone provides an important transition to the less resilient subchondral bone. For a long time this was regarded more or less as an inactive zone, until Hunziker (1992)7 noted that also the chondrocytes here could take up and incorporate (35S) sulfate into the pericellular and territorial matrix. Hunziker speculated that, following trauma, the metabolic activity here becomes temporarily impaired.7
Regarding experimental animals, it is important to know that it is only in adult animals that the division into zone I to zone III is possible.8 In the immature animal, the cells are more randomly distributed with a gradient in cell size from the top to the calcified zone, with the cells in the deeper parts being largest. Thus, the articular cartilage organization during prepubertal growth imitates the structure of the growth plate, and during that time the biomechanical properties of the cartilage change with an increase in stiffness and in shearing and compressive resistance.7,9
Metabolic Events in the Cartilage
The majority of the proteoglycans have a life span of about 600 days, but a small proportion of the proteoglycans in adult cartilage act as a fast fraction with a half-life of about 8 days. The proteoglycans are thus also more vulnerable to enzymatic degradation.10,11
This is a sophisticated and well-balanced process regulated by the chondrocytes, and a disturbance of any of these events could lead to destruction of the cartilaginous matrix. This happens in osteoarthritis where an early sign is an imbalance in synthesis and degradation of the matrix.
Related posts:
 Postoperative Cartilage Repair Rehabilitation
Postoperative Cartilage Repair Rehabilitation
 Bone Grafting around an Articular Joint
Bone Grafting around an Articular Joint
 Autologous Chondrocyte Implantation: ACI First and Second Generation
Autologous Chondrocyte Implantation: ACI First and Second Generation
 Bone Marrow Stimulating Techniques: Carbon Fiber Resurfacing
Bone Marrow Stimulating Techniques: Carbon Fiber Resurfacing
 Fixation of Osteochondral Fragments
Fixation of Osteochondral Fragments
 Autologous Chondrocyte Implantation: Transarthroscopic Implantation of Hyalograft (Hyaff 11) with Autologous Chondrocytes
Autologous Chondrocyte Implantation: Transarthroscopic Implantation of Hyalograft (Hyaff 11) with Autologous Chondrocytes



