CHAPTER 3 Anatomy and physiology of skin and soft tissue
1. Describe the mechanisms by which the skin is able to provide its six major functions.
2. Describe the structures, function, and cellular composition of the skin layers, including layers within the dermis and epidermis.
3. Explain the relationship between skin pigmentation and protection against ultraviolet radiation.
4. Compare and contrast the structural and cellular development of the skin in the fetus, premature infant (between 23 to 32 weeks’ gestation), full-term neonate, adolescent, and adult.
5. Describe at least two effects on the skin of the following: hydration, sun, nutrition, soaps, and medications.
6. Compare and contrast the structure and function of dark and lightly pigmented skin.
Population projections indicate that changing demographics will occur over the next decades, and that by 2042 a majority of the US population will be composed of people with skin of color: Hispanic (30%), African American (15%), Asian (9.2%), American Indian and Native Alaskan (2%), and Native Hawaiian and other Pacific Islanders (0.6%) (US Census Bureau, 2008). Unique characteristics of darker versus lighter pigmented skin (Box 3-1) are described throughout the chapter.
BOX 3-1 Unique Characteristics of Darker Pigmented Skin
• Higher lipid content in stratum corneum
• Increased junctional integrity
• Stratum corneum contains melanosomes (light skin has no melanosomes in stratum corneum)
• Inflammatory reactions (i.e., reddened areas) can be masked
• Inflammation may appear violet-black or black
• Vitiligo and keloids more common
• More blood vessels and dilated lymph channels
• Less abundant distribution of elastic fibers
• Greater protection against photoaging or dermatoheliosis due to no obvious signs of damage to elastic fibers upon exposure to sun light
• Increase in hair breakage and higher prevalence of traumatic alopecia
• More prone to pseudofolliculitis due to spiral configuration and curvature of hair follicles
• Better protection against skin cancer due to increased synthesis, amount, distribution of melanin
• Can mask precancerous and cancerous tumors due to increased melanin content
Skin layers
Human skin is divided into two primary layers: epidermis (outermost layer) and dermis (innermost layer) (Figure 3-1). These two layers are separated by a structure called the basement membrane. Beneath the dermis is a layer of loose connective tissue called the hypodermis, or subcutis.
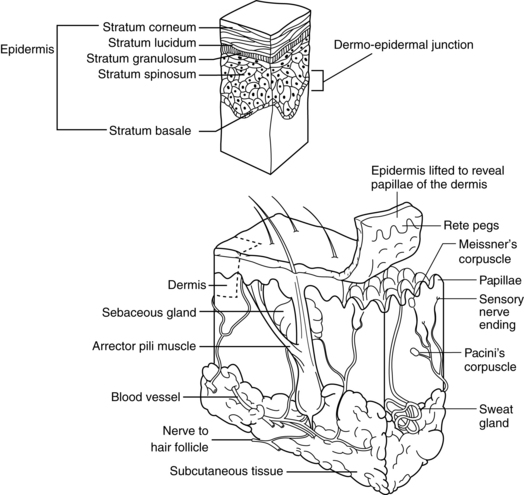
FIGURE 3-1 Schematic diagram of anatomy of skin and subcutaneous tissue.
(From Hooper BJ, Goldman MP: Primary dermatologic care, St. Louis, 1999, Mosby.)
Epidermis
The epidermis, the outermost skin layer, is avascular and derived from embryonic ectoderm. This layer is relatively uniform in thickness over the body, between 75 and 150 μm, except on the soles and palms, where thickness is between 0.4 and 0.6 mm. The epidermal layer is constantly being renewed, with turnover time ranging from 26 to 42 days. Complete epidermal renewal occurs over a period lasting between 45 and 75 days, or about every 2 months (Odland, 1991). The epidermal layer is composed of stratified squamous epithelial cells, or keratinocytes, and is divided into five layers (see Figure 3-1). These layers, beginning from the outermost to the innermost, are the stratum corneum, stratum lucidum, stratum granulosum, stratum spinosum, and stratum basale (stratum germinativum or, simply, the basal layer).
Stratum corneum.
The stratum corneum, or horny layer, is the top layer and is composed of dead keratinized cells. These squames, or corneocytes, are the cells that are abraded by the daily mechanical and chemical trauma of handwashing, scratching, bathing, exercising, and changing of clothes. The stratum corneum is composed of layers of thin, stacked, pancake-appearing, anucleate cells. Approximately 80% of these cells are filled with keratin, a tough, fibrous, insoluble protein; hence, they are called keratinocytes. Keratinocytes are initially formed in the basal layer and undergo the process of differentiation. The normal stratum corneum is composed of completely differentiated keratinocytes. Keratin is resistant to changes in temperature and pH and to chemical digestion by trypsin and pepsin. This same protein is found in hair and nails; in these structures, keratin is referred to as “hard” keratin compared with the “soft” keratin of the skin (Jacob et al, 1982; Solomons, 1983). Stratum corneum thickness varies with age, gender, and disease. In the deeper layer of the stratum corneum, the stratum compactum, the keratin is more densely packed and the cells have a diminished capacity to bind water (Haake, Scott, and Holbrook, 2001). In the upper layer, the stratum dysjunctum, cells are partly shed as a result of proteolytic degradation of the desmosomes.
The stratum corneum is enriched with a lipid matrix that enhances the barrier properties of the skin. Although not universally agreed upon, across the skin color spectrum, differences in the stratum corneum exist as summarized by Taylor (2002). At least two studies (LaRuche and Cesarini, 1992; Rienertson and Wheatley, 1959) report that black stratum corneum has a higher lipid content and this may in part contribute to the conflicting results regarding strateum corneum thickness. The stratum corneum in darkly pigmented skin has been reported to have increased junctional integrity as evidenced by the increased number of tape strippings required to remove the stratum corneum. Whether the stratum corneum is thicker in darkly pigmented skin is unclear (Taylor, 2002).
Stratum lucidum.
The stratum lucidum is directly below the stratum corneum. This layer is found in areas where the epidermis is thicker, such as the palms of the hands and the soles of the feet, but it is absent from thinner skin, such as the eyelids. This layer can be one to five cells thick and is transparent. Cell boundaries often are difficult to identify in histologic sections under a light microscope. The stratum lucidum is a transitional layer where active lysosomal enzymes degrade the nucleus and cellular organelles before they are moved into the stratum corneum (Jacob et al, 1982; Wysocki, 1995).
Stratum granulosum.
The stratum granulosum, or granular layer, is beneath the stratum lucidum when the stratum lucidum is present; otherwise, the stratum granulosum lies beneath the stratum corneum. This layer is one to five cells thick and is so named because of the granules present in the keratinocytes of this layer. The cells of the stratum granulosum have not yet been compressed into a flattened layer and are diamond shaped. The structures contained in these cells are keratohyalin granules, which become intensely stained with the appropriate acid and basic dyes. The proteins contained in these granules (profilaggrin, intermediate keratin filaments, and loricrin) help to organize the keratin filaments in the intracellular space. Profillaggrin is cleaved to form filaggrin, which is further degraded to form urocanic acid and pyrrolidone carboxylic acid. Both urocanic acid and pyrrolidone carboxylic acid provide additional hydration to the stratum corneum and limited filtering against ultraviolet radiation (UVR). The major component of the cornified envelope is loricrin, which is cross-linked to other protein components (involucrin, cystatin A, small proline-rich proteins [SPRR1, SPRR2], elafin, envoplakin) that together form up to 70% of the molecular mass of the cells in this layer. Cells in this layer still have active nuclei (Millington and Wilkinson, 1983; Wheater et al, 1987).
Stratum spinosum.
The stratum spinosum is below the stratum granulosum. This layer often is described as the prickly layer because cytoplasmic structures in these cells take on this morphology. Generally the cells of this layer are polyhedral. A prominent feature of the prickle layer is the desmosome, a type of cell–cell junction. The desmosomes provide adhesion between cells and resistance to mechanical forces. Cells in this layer begin to synthesize involucrin, a soluble precursor of the cornified envelopes (Millington and Wilkinson, 1983). The spinous cells contain large bundles of newly synthesized keratin filaments (K1/K10) in addition to K5/K14 still present from the basal layer (Haake, Scott, and Holbrook, 2001). The identification of various classes of desmosomal proteins (plakoglobin, desmoplakin, plakophilins) and transmembrane glycoproteins (desmogleins, desmocollins) have led to the identification of their roles in various epidermal pathologies, such as bullous impetigo, pemphigus vulgaris, and other genodermatoses.
Stratum basale or stratum germinativum.
The stratum basale, or stratum germinativum, is the innermost epidermal layer. It often is referred to simply as the basal layer (see Figure 3-1). It is a single layer of mitotically active cells called basal keratinocytes, or basal cells. These active cells respond to several factors, such as extracellular matrix, growth factors, hormones, and vitamins. Skin metabolism is mediated by glucose. Glucose utilization in the skin is comparable to that in muscle. Glucose leaving the circulation crosses the basement membrane and forms a concentration gradient that decreases as the glucose moves to the upper layers of the epidermis.
Once cells leave the basal layer they begin an upward migration, which can take 2 to 3 weeks. A cell takes approximately 14 days to move to the stratum corneum and another 14 days to move through the stratum corneum and desquamate (Haake, Scott, and Holbrook, 2001). After leaving the basal layer, the cells begin the process of differentiation. All layers of the epidermis consist of peaks and valleys, but this arrangement is more dramatic in the basal layer such that these protrusions are partly responsible for anchoring the epidermis, thus providing structural integrity. These epidermal protrusions of the basal layer point downward into the dermis and are called rete ridges, or rete pegs. The basal layer is the primary location of mitotically active cells of the epidermis and has been noted to have increased proliferative capacity compared with cells at the top of the ridges (Briggamann, 1982).
Epidermal stem cells compose about 10% of the basal cell population. These stem cells have a slow cell cycle. When labeled with a radiolabeled DNA precursor, epidermal stem cells retain the label for long periods, thus they are identified as label-retaining cells (Bickenbach 1981). Once cell division occurs, daughter cells that will undergo differentiation as they move toward the stratum corneum are called transient amplifying cells. These transient amplifying cells make up approximately 50% of the basal keratinocyte population. Transient amplifying cells progress to become postmitotic cells that then terminally differentiate. A portion of the stem cell population is found in the epidermal crypts of the rete pegs and in the bulge region of hair follicles. Dividing cells go through the cell cycle, and the G1 phase is shortened in states such as wound healing. The normal keratinocyte cell cycle time is 300 hours but may be as short as 36 hours when psoriasis is present. These stem cell compartments and the transient amplifying cells both are capable of limited or continuing cell division and thus are the cells most likely to reside long enough in the skin to undergo genetic modifications that lead to the development of skin cancers. The use of stem cells alone and in combination with tissue engineering approaches is an area of active research and development in wound care (Griffith and Naughton, 2002).
Melanocytes, the cells responsible for skin pigmentation as a result of melanin synthesis, also are distributed in the basal layer. Melanocytes are dendritic cells that arise from melanoblasts, which derive from the neural crest. During development, they migrate to other locations, including the bulge region of hair follicles, the choroid of the eye, the heart, and the brain (Goding, 2007). Melanocytes are also required for hearing; genetic mutations resulting in a loss of melanocytes are a major cause of deafness (Steel and Barkway, 1989). Melanin is packaged inside the cell into melanosomes, which are transported via the cytoskeletal network of microfilaments and microtubules to the dendritic processes of the melanocyte, where they are then transferred to keratinocytes. Incorporation into the keratinocytes is thought to occur via one or more of the following processes: release and endocytosis, engulfment, active transport, or passage through channels or pores between the melanocyte and neighboring keratinocytes (Boissy, 2003). Melanocytes can be detected by 50 days’ gestation in fetal development (Holbrook, 1998). Under normal conditions, melanocytes rarely divide. In normal skin, the number of melanocytes present is nearly the same regardless of skin color. There is approximately one melanocyte for every 36 basal cells. Dendritic melanocyte structures are responsible for the transfer of pigment to a large number of keratinocytes. The primary difference between light- and dark-skinned individuals is the size, number, and distribution of melanosomes, the structures containing the melanin pigment, and the activity of the melanocytes. Carotene or carotenoids are responsible for imparting the yellow hue to the skin of some individuals (Jacob et al, 1982; Sams, 1990; Solomons, 1983).
Melanin pigment is a complex polymer that is synthesized from tyrosine by activity of the tyrosinase gene pathway. Melanin is difficult to study because it is relatively insoluble (Boissy, 2003). The two primary types of melanin are brown/black eumelanin and red/yellow pheomelanin (Lin and Fisher, 2007). Pheomelanin is more photolabile, and damage leads to release of oxygen radicals such as hydrogen peroxide, superoxide, and hydroxyl radicals, resulting in oxidative stress and DNA damage (Lin and Fisher, 2007). Constitutive synthesis, or the synthesis that occurs at basal levels, results primarily from genetic regulation. Facultative synthesis is the additional up- or down-regulation of melanin synthesis in response to UVR, hormones, cytokines, immune regulation, chemical exposure, or other agents (Boissy, 2003). Interestingly, once delivered to the keratinocytes, melanosomes are preferentially, although not exclusively, clustered around the apical side of the nucleus, where they can more effectively provide protection from UVR to prevent DNA damage leading to mutations (Boissy, 2003). In addition to its photoprotective effect, melanin is thought to act as a sink for highly reactive oxygen species that can also lead to DNA damage (Goding, 2007). Pathologies associated with melanocytes or pigmentation are melanoma, dyschromias such as vitiligo and melasma, xeroderma pigmentosa, and albinism.
As a result of genetics, genetic recombination, and selective environmental pressures (e.g., UVR intensity and exposure, age, hormonal influences), skin pigmentation demonstrates considerable variation across the human population. Presently more than 150 genes have been identified that either directly or indirectly affect skin color (Yamaguchi and Hearing, 2009). Key proteins and receptors identified with the synthesis and transport of melanin or melanosomes include melanocyte-stimulating hormone, melanocortin-1 receptor (MC1R), keratinocyte growth factor, and protease-activated receptor-2. The red hair/light skin phenotype is associated with mutations found in the residues of MC1R, which lead to increased risk for skin cancer (Miyamura et al, 2006). Thus, the skin color spectrum ranges from dense black, with the highest level and distribution of melanosomes, to various shades of brown to white and finally to albino, or the complete absence of pigment (but not an absence of melanocytes). In general, Caucasian and Asian skin has smaller melanosomes that form small clusters or aggregates, whereas dense African skin has larger melanosomes that are distributed primarily as single units (Montagna et al, 1993; Szabo et al, 1969; Szabo et al, 1972). In darkly pigmented skin, despite the hydrolytic degradation that occurs with keratinocyte differentiation, some melanosomes can still be seen in the stratum corneum, whereas in light skin no melanosomes are seen in the stratum corneum (Boissy, 2003). Various shades of brown skin occur across African, African American, Hispanic, Asian, Caucasian, and Native American peoples; these shades are influenced by the number, size, and distribution of melanosomes. The range of variation can even occur in the same individual, depending on skin location and sun exposure (Szabo, 1954). In general, the density of melanocytes is slightly higher in skin covering the upper dorsal surfaces compared to skin on the lower dorsal surfaces, and the amount of melanin present in the skin correlates well with visible skin pigmentation seen on physical examination (Miyamura et al, 2006).
Intensely pigmented skin can mask the detection of skin inflammatory reactions, which normally are seen as reddened areas that may appear in response to conditions such as contact dermatitis, pressure, or folliculitis. Inflammation may appear as darker areas in black skin, violet-black in intensely black skin, or black in brown skin. Other signs of inflammation may include the detection of heat or warmth to touch or induration detected as skin tightening or hardening over areas where there is skin damage. Dermatologic disorders accompanied by inflammation can result in postinflammatory hypopigmentation or hyperpigmentation that can be especially distressing to individuals with more darkly pigmented skin in which the color contrast can be dramatically evident. Other dermatologic disorders more commonly associated with darkly pigmented skin include vitiligo and keloids. Keloids occur 3 to 18 times more frequently in darkly pigmented skin, and the incidence has been reported as between 4.5% to 16% in Chinese, Hispanic, and African American individuals (Oluwasanmi, 1974; Robles and Berg, 2007; Taylor, 2002; Taylor, 2003).
Universal agreement concerning comparative differences, if any, across the skin color spectrum for transepidermal water loss (TEWL), strateum corneum structure, epidermal thickness, and sweat and sebaceous gland number or distribution, are fraught with numerous points of criticism that include low numbers of subjects, variation in age, sex, circadian rhythms, use of emollients, relative temperature and humidity, geographic location, location of skin sample, sun exposed versus sun protected areas, nutrition, validity and reliability of instrumentation, and accurate indexing to skin color (McDonald, 1988; Taylor, 2002). It has been noted that there are 31 climate zones around the globe subject to the adaptation, lifestyle, and conditions of people with various customs and resources and climate can influence birth weight, body shape, and cranial morphology (Lambert et al, 2008), Thus, definitive studies of skin biology across the range of the skin color spectrum relative to age and gender await more exhaustive and comprehensive research studies and may require more precise instrumentation for some measures.
Basement membrane zone
The basement membrane zone (BMZ), or dermal–epidermal junction, is the area that separates the epidermis from the dermis. Closer examination of the BMZ in the past decade has revealed the BMZ to be more complex than previously believed. Basal keratinocytes use hemidesmosomes to structurally and functionally attach to the BMZ. The BMZ is subdivided into two distinct zones, the lamina lucida and the lamina densa. The lamina lucida is so named because it is an electron-translucent zone compared with the electron-dense zone of the lamina densa. The major proteins found in the BMZ are fibronectin, an adhesive glycoprotein; laminin, a glycoprotein; type IV collagen, a non–fiber-forming collagen; and heparin sulfate proteoglycan, a glycosaminoglycan. A lesser amount of type VII collagen has also been detected. The BMZ anchors the epidermis to the dermis and is the layer that is affected in blister formation associated with dermatologic diseases, second-degree burns, full-thickness wounds, and mechanical trauma. During wound healing the BMZ is disrupted and must be re-formed (Sams, 1990).
Dermis
The major proteins found in the dermis are collagen and elastin. The other category of proteins found occupying the space between collagen and elastin fibers is referred to as the ground substance. The ground substance in black skin is distinctly more abundant in both the papillary and reticular layers than in white skin (Montagna and Carlisle, 1991). This category of proteins is largely composed of proteoglycans and glycosaminoglycans. Included in this category of proteins are chondroitin sulfates and dermatan sulfate (versican, decorin, biglycan), heparan and heparan sulfate proteoglycans (syndecan, perlecan), and chondroitin-6 sulfate proteoglycans. Although these proteins account for only approximately 0.2% of the dry weight of the dermis, these large molecules are capable of binding up to 1,000 times their volume. Thus, proteoglycans and glycosaminoglycans play a role in regulating the water-binding capacity of the dermis, which can determine dermal volume and compressibility. Hyaluronan is also found in the dermis and in higher abundance in fetal skin, where it forms a watery, less stable matrix that allows greater cell movement; it also is a critical factor in scarless healing (Longaker et al, 1990). Hyaluronan can bind growth factors and provide cellular linkages with other matrix materials that together function to regulate cell migration, adhesion, proliferation, differentiation, morphogenesis, and tissue repair (Haake and Holbrook, 1999). Other glycoproteins found in the dermis include fibronectin, thrombospondin, laminin, vitronectin, and tenascin. These glycoproteins are synthesized and secreted by fibroblasts in the dermis.
Data indicate that skin fibroblasts from different anatomic locations contain different transcriptional patterns that can indicate position and can be differentiated from fibroblasts in other locations using microarray analysis (Chang et al, 2002). Fibroblasts present in the dermis range in size and number. It has been reported that fibroblasts in black facial skin were larger, occurred in greater numbers, and were more likely to be binucleated or multinucleated compared to white skin. Overall, the dermis in black skin was thick and compact compared to thinner and less compact in white skin (Montagna and Carlisle, 1991). In this same study, collagen fiber bundles were observed to be smaller, more tightly packed and were oriented parallel to the epidermis in black skin and had more fiber fragments. Somewhat in contrast, white skin was noted to have larger collagen fiber bundles with some degraded collagen fragments, which may have resulted from sun exposure, but had greater variability in the number of fibroblasts. No differences in mast cells were observed. However, black skin did have more binucleated and multinucleated macrophages in the papillary dermis, and the accompanying melanophages in black skin contained melanosomes both singly and in complexes, whereas in white skin the melanophages were almost exclusively in complexes (Montagna and Carlisle, 1991). Again, these findings should be viewed with caution given the lack of comprehensive studies across the skin color spectrum. The dermis is a matrix that supports the epidermis. It can be divided into two areas: papillary dermis and reticular dermis (see Figure 3-1). The papillary and reticular layers appear more distinct in white skin and less distinct in black skin (Montagna and Carlisle, 1991).
Papillary dermis.
The papillary dermis lies immediately below the BMZ and forms interdigitating structures with the rete ridges of the epidermis called dermal papillae. The dermal papillae contain papillary loops (Figure 3-2), which supply the necessary oxygen and nutrients to the overlying epidermis via the BMZ. The collagen fibers contained in the papillary dermis are much smaller in diameter and form smaller, wavy, cablelike structures compared with the reticular dermis. This portion of the dermis also contains small elastic fibers and has a greater proportion of ground substance than the reticular dermis. The superficial subepidermis in darkly pigmented skin contains more numerous blood vessels along with many more dilated lymph channels (Montagna and Carlisle, 1991). The papillary dermis also contains lymphatic vessels that play a role in controlling interstitial fluid pressure by resorption of fluid and assist in clearing the tissues of cells, lipids, bacteria, proteins, and other degraded substances. The papillary lymph vessels drain into the horizontal plexus of larger lymph vessels located in the deep subpapillary venous plexus. This system is further connected to the lymph system in the reticular dermis. Fluid flow through this system is partially controlled through arterial pulsations, muscle contractions, and body movement. Renewed interest in the skin lymphatic system has resulted from the identification of markers such as VEGF-C, VEGFR-3, and lymphatic vessel endothelial receptor-1 (LYVE-1) and the findings of the role of these lymphatic vessels in tumor promotion (Chu, 2008).
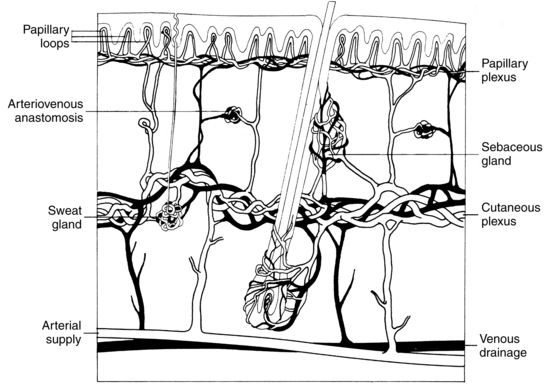
Dermal proteins.
Collagen, the protein that gives the skin its tensile strength, is the major structural protein found in the dermis and accounts for 25% of the skin’s total weight (Stryer, 1995). The primary constituents of collagen are proline, glycine, hydroxyproline, and hydroxylysine. Collagen is secreted by dermal fibroblasts as tropocollagen, which undergoes additional extracellular processing so that mature collagen fibers are formed. Normal human dermis is primarily composed of type I collagen, a fiber-forming collagen. Type I collagen represents 77% to 85% of the collagen present; type III collagen, also a fiber-forming collagen, represents the remaining 15% to 23% (Gay and Miller, 1978; Wysocki et al, 2005). Very small amounts of type V collagen (less than 5%) and type VI collagen also are present.
Elastin, another protein found in the dermis, provides the skin’s elastic recoil, a feature that prevents the skin from being permanently reshaped. It is a fiber-forming protein, like collagen. Elastin has a high amount of proline and glycine. However, unlike collagen, elastin lacks large amounts of hydroxyproline. Elastin fibers form structures, similar to coils, that allow this protein to be stretched and, when released, to return to its inherent configuration. Elastin accounts for less than 2% of the skin’s dry weight (Millington and Wilkinson, 1983; Sams, 1990; Wysocki, 1995). Elastin is distributed with collagen but in smaller amounts. Large concentrations of elastin are found in blood vessels (especially the aortic arch near the heart) and lymphatic vessels. The distribution of elastic fibers ranges from less abundant in darkly pigmented skin to more abundant in light skin. Upon exposure to sunlight, elastic fibers in darkly pigmented skin show no obvious signs of damage, whereas elastic fibers in light skin show obvious signs of elastosis (Montagna et al, 1993). The lower distribution of elastic fibers in darkly pigmented skin also extends to the anchorage of hair follicles, leading to an increase in hair breakage and a higher prevalence of traumatic alopecia in both men and women (Taylor, 2002). Furthermore, the curvature of hair follicles and the spiral configuration seen in Africans and African Americans lead to pseudofolliculitis, especially in men who shave.
Hypodermis
Hypodermis, or superficial fascia, forms a subcutaneous layer below the dermis. It is an adipose layer containing a subdermal plexus of blood vessels giving rise to the cutaneous plexus in the dermis, which in turn gives rise to the papillary plexus and loops of the papillary dermis (see Figure 3-2). The hypodermis attaches the dermis to underlying structures. This layer provides insulation for the body, a ready reserve of energy, and additional cushioning; it also adds to the mobility of the skin over underlying structures (Haake and Holbrook, 1999). Growing hair follicles and apocrine and eccrine sweat glands can extend into this layer. The hypodermis is largely absent in certain pathologic disease states, such as Werner syndrome and scleroderma. Adipocytes form the primary cells in this tissue layer, and their activity is regulated by leptin.
Skin functions
The skin provides several functions: protection, immunity, thermoregulation, sensation, metabolism, and communication (Jacob et al, 1982; Millington and Wilkinson, 1983; Woodburne and Burkel, 1988).
Protection
The skin protects the body against aqueous, chemical, and mechanical assaults; bacterial and viral pathogens; and UVR. It also prevents excessive loss of fluids and electrolytes in order to maintain the homeostatic environment. The effectiveness of the skin in preventing excessive fluid loss can be seen in patients with burns. Patients with burns involving 30% of their body can lose up to 4.1 L of fluid compared with 710 ml for normal adults (Rudowski, 1976). The skin maintains the internal milieu, and a progressive decrease in water content across the epidermal layers helps to prevent excessive transepidermal water loss under basal conditions. Water constitutes 65% to 70% of the basal layer, and decreases to 40% in the granular layer and 15% in the stratum corneum (Warner et al, 1988). The skin’s barrier function is so effective that percutaneous drug delivery is limited except in premature babies and in newborns. In general, compounds are restricted to those with a molecular mass of 500 daltons or less with a daily dose of 10 mg or less. Protection against mechanical assaults is mainly provided by the tough fibroelastic tissue of the dermis, collagen, and elastin.
Protection against pathogens.
Sebum, a lipid-rich, oily substance secreted by the sebaceous glands onto the skin surface, usually via hair follicles and shafts, provides an acidic coating with a pH ranging from 4 to 6.8 (Spince and Mason, 1987) and a mean pH of 5.5 (Roth and James, 1988; Wysocki et al, 2005). This acidity, together with natural antibacterial substances found in sebum, retards the growth of microorganisms. These glands are stimulated by sex hormones (androgens) and become very active during adolescence. Sebum, along with keratin, provides resistance to aqueous and chemical solutions. When sebaceous glands occur in association with hair follicles, they are called a pilosebaceous unit. Sebaceous glands are not found on palms or soles; they occur in areas that lack hair, such as lips. These glands are largest when they are located on the face and when associated with hair follicles. Sebaceous glands may increase in size by 100 to 150 times as sebum accumulates. Maximum secretion occurs in persons in their late teens to early twenties. Rates of secretion are higher in men and decline 32% per decade in females compared to 23% per decade in males.
Resistance to pathogenic microorganisms is also provided by normal skin flora through bacterial interference (Noble, 2004; Weinberg and Swartz, 1987). Conceptually, the two categories of skin flora are (1) resident (bacteria normally found on a person) and (2) transient (bacteria not normally found on a person and usually shed by daily hygienic practices, such as bathing and hand washing). Resident bacteria are found on exposed skin; moist areas such as the axillae, perineum, and toe webs; and covered skin. Bacterial microcolonies are found in hair follicles and at the edges of squames as halos in the upper loose surface layers. The following species of bacteria are found on human skin: Staphylococcus, Micrococcus, Peptococcus, Corynebacterium, Brevibacterium, Propionibacterium, Streptococcus, Neisseria, and Acinetobacter. The yeast Pityrosporum and the mite Demodex are also found. Not all species are found on any one individual, but most individuals have at least five of these genera. Normal viral florae are not known to exist (Noble, 2004). However, viruses have been detected in compromised skin and in individuals who are not immunocompetent.
An association exists between skin pH and bacteria. For example, Propionibacterium acnes grows well at pH values of 6 and 6.5, but its growth is markedly decreased at pH 5.5 (Korting and Braun-Falco, 1996). However, this association can vary depending on the specific bacterial species involved and the body location. The formation of bacterial biofilms has been scrutinized (Wysocki, 2002), and the role of normal skin flora in the formation of these biofilms is an area of active investigation. Biofilms are essentially an extracellular polysaccharide matrix, or glycocalyx, in which microorganisms are embedded. Biofilms are composed of mixed bacterial species living in their own microniche in a complex, metabolically cooperative microbial community that maintains its own form of homeostasis and rudimentary circulatory system (Costerton et al, 1995). Biofilms are resistant to host immune responses and are markedly more resistant to antibiotic and topical bactericidals (Xu et al, 2000). Reports indicate that biofilm cells can be at least 500 times more resistant to antibacterial agents (Costerton et al, 1995). Biofilm formation is familiar to clinicians and can be identified on endotracheal tubes, Hickman catheters, central venous catheters, contact lenses, and orthopedic devices. Pseudomonas aeruginosa forms biofilms in conjunction with other bacterial species. Quorum sensing by bacteria is a feature in biofilm formation. These biofilms complicate the eradication of infections because they give rise to sessile and planktonic bacteria. Planktonic bacteria can be cleared by phagocytosis, antibodies, and antibiotics (Costerton et al, 1999). Sessile bacteria in biofilms can evade antibiotics by giving rise to planktonic bacteria, which respond to antibiotics and host immune responses, but the sessile bacteria remain. Cycles of antibiotic treatment often are administered without success, and the symptoms of infection recur. In these situations, biofilm formation should be suspected, and surgical removal of the sessile population most likely will be required to eliminate the pathogenic bacteria. Moist skin areas, such as the axillae, perineum, toe webs, hair follicles, nail beds, and sweat glands, are especially prone to an increased presence of bacteria. Protection from bacterial invasion is mediated in part by proteins called defensins (Hoffman et al, 1999). Findings show that β-defensin is abundant in skin and may be important for wound healing. Defensins act in conjunction with phagocytosing neutrophils and the membrane attack complex of complement. Thus, optimal skin and wound care is a cornerstone of clinical practice in preventing the progression from bacterial colonization to infection (Wysocki, 2002).

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


