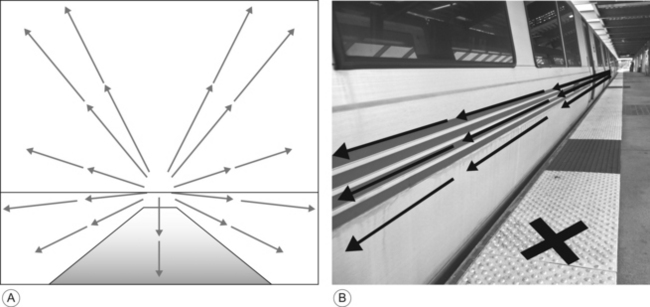
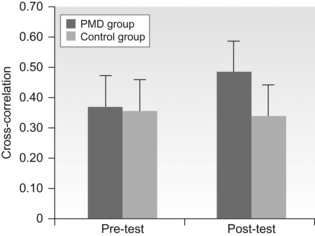
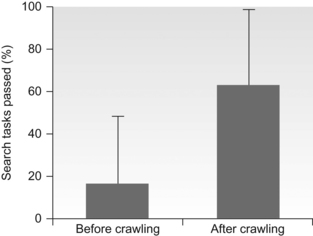
10 David I. Anderson, Joseph J. Campos, Monica Rivera, Audun Dahl, Ichiro Uchiyama and Marianne Barbu-Roth Brief historical perspective on motor activity and psychological functioning Independent mobility and psychological development in the human infant Mobility and psychological development: implications for infants with disabilities Conclusions and future directions The field of developmental paediatrics has long held a bias against motoric processes affecting psychological development (Young, 1977). Related to this bias is the established paradigm for analysing children with developmental disabilities—a paradigm wherein causal analysis begins with the brain and stops at the musculature, failing to consider how the process in fact continues and loops back from movement to brain. This chapter challenges these biases and presents evidence from a multitude of carefully crafted experiments documenting that motoric developments bring about profound psychological reorganizations in an array of crucial psychological domains. We maintain that these motorically paced reorganizations are not well known, or at all, by segments of the paediatric community; and we propose implications for understanding cerebral palsy (CP) and similar motoric disabilities, the functional deficits in whom are thus too often not appreciated. This chapter concentrates on the effects of self-produced locomotion on changes in spatial-cognitive functioning and the implications these effects have for children with physical disabilities like CP. Successful independent mobility is an excellent example of the more general effects that motor activity can have on psychological development. We believe that the early promotion of functional mobility is crucial in maximizing children’s potential for independent living. Our contribution is in showing that independent mobility has consequences that extend far beyond independence in everyday activities. Mobility generates and maintains psychological changes that are crucial for adapting to a complex world (Campos et al., 2000). This idea, which is an illustration of the well-accepted notion that animals play an active role in their own development, is not new. However, its translation to the clinical setting has lagged behind the experimental discoveries (Campos et al., 1982), despite the fact that many children with disabilities could profit from the knowledge at hand. Modern philosophers and scientists have speculated about movement’s role in perception and cognition since the 1700s, when Bishop Berkeley published An Essay Towards a New Theory of Vision. Berkeley argued that because visual stimuli were impoverished and ambiguous, motor experience was necessary to make sense of the visual world. His ideas inspired a number of different motor theories of perception that were popular at various times during the 20th century (for a review, see Weimer, 1977). The 1950s were a particularly generative period for theorizing about the role of motor activity in psychological functioning. Sherrington (1951) highlighted that the brain evolved to control movement and so movement must be fundamental to thinking. Sperry (1952) argued that the entire output of our thinking consists of little more than patterns of movement and Piaget (1952, 1954) stressed that the origins of intelligence were in an infant’s sensorimotor interactions with the world. Like Berkeley, Piaget believed that motor experience was necessary to structure patterns of sensory stimuli that were initially meaningless. The theorizing in the 1950s set the stage for a wave of important animal experiments in the late 1950s and 1960s. Those experiments showed convincingly that the normal development of visual–motor co-ordination was dependent on self-produced movement. Held and Hein’s (1963) kitten experiment is the best-known illustration of the early plasticity of the perceptual systems and the critical link between movement experience and perceptual development. Kittens were raised in the dark and then exposed to a patterned environment in an active or passive locomotion condition. All kittens moved along the inside perimeter of a black and white striped cylinder. However, the passive kittens rode in a gondola that was yoked to the movements of the active kitten via a horizontal rod and tether. After training, only the active kittens showed spatially adaptive behaviour by avoiding a drop-off and extending their forelimbs in preparation for contact with a surface. In a clever confirmation and extension of the original findings, Hein et al. (1970) covered one of the kitten’s eyes with a patch as it moved actively in the cylinder. The kittens behaved adaptively in response to the drop-off and surface only when tested with the active eye but not the passive eye, highlighting the specificity of the effects of locomotor experience on visual–spatial development. Though widely cited in the psychological literature, Held and Hein’s work did not stimulate a wave of research on the motor contributions to perceptual and psychological development that might have been expected. On the contrary, the role of motoric factors in psychological development is still downplayed by the scientific community, despite an impressive body of empirical evidence suggesting that motor activity makes a central contribution to psychological function (Campos et al., 2000; Rakison and Woodward, 2008). Bertenthal and Campos (1990) listed three reasons why the study of the functional consequences of motor development had received little attention until then. The first was the apparent failure to confirm Gesell’s (1928) proposal that precocious motor development in infancy predicts IQ in later childhood. The second was the lack of methods available until recently to adequately measure motor processes, and the third was the tendency to explain developmental changes on the basis of innate predetermined mechanisms (e.g., Diamond, 1990), a bias that continues to exist in many circles (e.g., Spelke and Kinzler, 2009). The limitations of quasi-experimental designs have further encouraged researchers to downplay the role of motor factors in psychological development because these designs cannot rule out maturation as an explanation for differences in psychological performance between groups that differ in motor competence. In fact, researchers consider the study of functional consequences of the causal link between motor development and cognitive (as well as social and emotional) development to be a methodological Gordian Knot. (In Greek mythology, the Gordian Knot was an intricate knot that resisted all attempts to untie it until Alexander the Great cut it with his sword. The Gordian Knot is often used as a metaphor for an intractable problem.) Haith and Benson (1998) commented: ‘It is not possible in any single study to control all key variables and potential confounds, including age of locomotor onset, age at spatial-cognitive testing, duration of locomotor experience, manipulation of type of locomotor experience, and level of locomotor skill. Researchers will need to account for these and other variables through multiple studies that converge on similar outcomes (p. 217).’ In subsequent sections we will show how our laboratory has addressed this challenge. Methodological issues are only part of the problem; the neglect of motor contributions to psychological development seems to be symptomatic of a broader neglect of the importance of motor processes in mainstream psychological phenomenon. In discussing the neglect, David Rosenbaum has gone as far as to dub motor control the Cinderella of psychology (Rosenbaum, 2005). Rosenbaum cites psychology’s intellectual roots in philosophy, where the contents of the mind have always held centre stage, as a primary reason for the tendency to overlook motor contributions to mental life. However, it is clear that the bias from philosophy cannot fully explain the neglect, given that many philosophers, including Berkeley, have shown a keen interest in the motor contributions to mind. In the words of the Bob Dylan, evidence suggests that ‘the times they are a-changin.’ Several trends have coalesced over the last few years to make the relation between motor activity and psychological function a legitimate and increasingly popular topic of investigation. The first trend has been the resurgence of interest in motor development, spurred largely by the popularity of the dynamic systems and the ecological approaches to the study of perception and action (Gibson and Pick, 2000; Thelen and Smith, 1994, 2006). Secondly, these approaches have also increased acceptance among psychologists of the idea that cognition is embodied; in other words, that our bodies and their potential for action provide the substrate for all psychological activity (Casasanto, 2011; Clark, 1997; Thelen, 2000; Varela et al., 1992; Wilson, 2002). Thirdly, researchers in the field of neuroscience, which has always had an interest in motor control, have discovered close anatomical linkages in brain areas known to be involved in cognitive and motor functions (e.g., Diamond, 2000). The linkage between the basal ganglia and the dorsolateral prefrontal cortex has been known for some time; however, Diamond (2000) has more recently argued that the connections between the cerebellum and the dorsolateral prefrontal cortex suggest that the cerebellum might also play an important role in cognitive functions. This suggestion is particularly relevant to cognitive deficits seen in children with CP because children with bilateral spastic CP are hypothesized to have disruptions to white matter tracts that connect the prefrontal and posterior brain regions, the basal ganglia, and related dopaminergic pathways, in addition to the disruptions to motor tracts (Bottcher, 2010; Christ et al., 2003). Some research suggests that the efficiency with which information is processed from the somatosensory system might also contribute to cognition. A study by Craft et al. (1995) showed that children with spastic diplegic CP who underwent selective dorsal rhizotomy to minimize overactive reflex activation in lower limb muscles showed dramatic improvements in attentional control and cognitive performance relative to controls who did not undergo the procedure. More recently, Dalvand et al. (2012) have supported this idea by arguing that the significant relation between motor and intellectual function seen in children with CP can be at least partially explained by a lack of mobility and consequently a lack of sensory stimulation to the brain regions involved in the control of motor and cognitive functions. The fourth trend relates to the most recent discoveries about brain plasticity. The role that experience plays in shaping the brain has garnered wide attention and at the same time has contributed to greater acceptance of the idea that motor activity influences brain development (Doidge, 2007; Gomes da Silva et al., 2012). Researchers are now more willing to accept a role for experience in developmental changes that were once considered genetically/maturationally determined. The fifth trend is the renewed interest shown in predicting academic performance from early measures of motor and cognitive function (Piek et al., 2008). Finally, the widespread problems of depression and obesity have popularized research into exercise/physical activity as non-pharmacological interventions (Ratey, 2007) and indirectly popularized examination of the broader link between motor activity and psychological function. Several studies have shown that exercise and physical activity induce the production of neurotransmitters and neurotrophins that are critical to learning, memory, and higher-order thinking (Erickson et al., 2012). Clinicians and researchers in the motor domain are well aware of the contribution earlier motor skills make to the development of later motor skills. Agency, the ability to act instrumentally to realize one’s goals or intentions, influences all aspects of development (Bidell and Fischer, 1997). Each new motor accomplishment heralds an increasing degree of independence and provides a foundation from which to launch the acquisition of new skills. Skill development occurs in a predictable sequence, such that each new skill builds logically on previous ones. For example, the onset of reliable head control and then sitting lead to step-function improvements in reaching, grasping, and manipulation (Rochat, 1992; Thelen and Spencer, 1998), and experience with belly crawling facilitates the development of proficiency in hands-and-knees crawling (Adolph et al., 1998). Many clinicians are unaware, however, that motor development contributes to developmental changes beyond the motor domain. Piaget (1952, 1954) built an entire theory of cognitive development on this premise, arguing that sensorimotor interactions with the world provide the building blocks for intellectual functioning. Piaget believed that sensory information is structured initially by the patterning of motor activity. Moreover, observing the consequences of movement permits the discovery of novel solutions to old problems and reveals new problems as the solutions to familiar goals are blocked by unexpected obstacles. An explosion of new goals, new choices, and potential interactions with the world confront infants as they become increasingly agentic. Agency recruits experience, which in turn builds agency. The two are interwoven throughout development in an intimate dance that permits an ever-increasing degree of autonomy and mastery over the environment. Although Piaget’s work has fallen out of favour, contemporary theorists have increasingly emphasized the importance of self-generated exploration as a driving force behind developmental change (Bidell and Fischer, 1997; Edelman, 1987; Gibson, 1988; Thelen and Smith, 1994). The link between independent mobility and psychological development is one of the best illustrations of the contributions that motor activity makes to psychological development. It is now clear that locomotor experience plays a central role in infant psychological development (Campos et al., 2000). Major changes in visual–motor co-ordination, visual–spatial understanding, problem solving, memory, emotion and social interaction occur after the onset of prone locomotion. The onset of prone locomotion has been associated more broadly with a reorganization in brain functioning (Bell and Fox, 1997). Thus, prone locomotion (typically in the form of hands-and-knees crawling) appears to catalyse a psychological revolution in the latter half of life’s first year, facilitating changes in an incredibly diverse set of skills that underlie adaptive behaviour. In this section, we will focus on changes in three areas—visual proprioception, spatial search, and spatial coding—to highlight how researchers have uncovered the relation between independent mobility and psychological development. These three areas were chosen because they index spatial cognition, a psychological skill that has attracted increasing attention in its own right because it provides a broad foundation for a host of higher-order intellectual skills that permit us to function in an increasingly technological society (Mix et al., 2010; Newcombe and Frick, 2010). James Gibson (1966, 1979) famously pointed out that vision is possible because of the structuring of ambient light reflected from the various surfaces that comprise the environment. Each surface has many tiny surfaces (patches or texture elements) that reflect bundles of narrow cones of light differently, depending on their orientation relative to the light source, their orientation relative to their neighbours, and their chemical composition. These bundles of reflected light form a densely structured optic array that is unique from any given point of observation. Consequently, when the head moves, whether through head turning, whole body postural sway, or locomotion, the surrounding optic array changes; it deforms or flows in a manner that co-varies precisely with the speed and direction of self-movement. Visual proprioception, then, is the optically produced awareness of self-motion that stems from the co-variation between self-motion and the optic flow field (Gibson, 1966, 1979; Lee, 1978, 1980). Optic flow is a rich source of information for the control of self-motion and its imposition on stationary observers can elicit very powerful postural and emotional reactions. Anyone who has felt their stomach drop while sitting or standing stationary in a simulated space flight or rollercoaster ride at an amusement park can attest to the powerful effect of optic flow. Visual proprioception is a widely under-appreciated psychological phenomenon, despite its obvious importance for the control of the postures and movements that are fundamental to the expression of all skilled activity (Anderson et al., 2004). The perception and control of self-motion is not only vitally important for motor skill development but also sensitivity to self-motion appears to play a central role in the development of the self—the sense of oneself as a unique entity, distinct from the external surround. The ability to perceive self-motion as distinct from motion of the surround provides the foundation from which the infant begins to differentiate herself from the environment (Neisser, 1988). This notion is particularly relevant to the current discussion, because recent research has shown that an infant’s ability to differentiate self-propelled from externally caused motion improves after the onset of crawling (Cicchino and Rakison, 2008). In addition, as we will see later, the ability to differentiate self-movement from externally generated movement has been implicated in a limited sense of agency experienced by children with CP (Ritterband-Rosenbaum et al., 2012). Although the ability to perceive and utilize optic flow has been examined relative to multiple aspects of behaviour (Gilmore et al., 2007; Warren and Wertheim, 1990), the primary focus here is on the development of visual proprioceptive control of head, trunk, and whole-body stability in sitting and standing. The developmental work on the visual proprioceptive control of balance has largely been conducted using the moving room paradigm, or a variant of the paradigm, originally designed by David Lee and colleagues (e.g., Lishman and Lee, 1973). The moving room is a large enclosure that can be moved back and forth relative to a stationary observer (see Fig. 10.1). Responsiveness to optic flow is assessed by measuring the infant’s postural sway in relation to movement of the walls. Figure 10.1 The moving room. The infant experiences global optic flow when all the walls are moved together, radial optic flow when only the front wall is moved, and lamellar optic flow when only the side walls and ceiling are moved. When looking in the direction of locomotion, the eyes are exposed to different geometrical patterns of optic flow; global optic flow in the entire field of view, radial optic flow (like a starburst pattern) in the central field of view, and lamellar optic flow (like the lines of longitude on a globe) in the periphery of the field (see Fig. 10.2). Stoffregen (1985) has observed that adults show greater postural responsiveness to lamellar optic flow in the periphery (generated by moving only the side walls of the room) than radial optic flow in the centre of the field of view (generated by moving only the front wall). Anyone who has experienced a false perception of self-motion when a train adjacent to the one in which you are sitting starts moving can appreciate the power of peripheral lamellar optic flow in conveying information about self-motion. Figure 10.2 (A) A pilot’s view of global optic flow experienced during landing an aircraft. The flow emanates from the location to which the pilot is heading. It has a radial structure in the centre of the field of view and an increasingly lamellar structure in the periphery. (B) Lamellar optic flow experienced when a train passes a passenger standing on the platform. When infants are exposed to global optic flow (whole room motion) in a moving room the magnitude of their postural compensation shows an interesting trajectory—it peaks during the early acquisition of a new posture and then diminishes as the posture comes under greater control only to peak again during the early acquisition of the next posture. For example, Butterworth and Pope (1981) have shown that infants as young as 2 months of age, when seated and provided with support for the trunk, will show postural compensation of the head when the whole room is moved toward them, with the magnitude of head sway decreasing with age and experience in controlling the head. Similarly, Butterworth and Hicks (1977) have observed that seated infants sway less in response to whole room movement as they gain increasing experience with the standing posture. However, the onset of standing and the onset of walking seem to be times when the infant is particularly responsive to global optic flow. The coupling between whole room motion and postural sway peaks between 9 and 10 months of age, at about the age where infants are just beginning to learn how to stand without support (though, curiously, also at the time they are mastering crawling) (Bertenthal et al., 1997; Delorme et al., 1989). Peaks in responsiveness to whole room movement that coincide with the onset of sitting, the onset of standing, and the onset of walking have been noted (Foster et al., 1996). Other research groups (Lee and Aronson, 1974; Schmuckler and Gibson, 1989; Stoffregen et al., 1987) have reported that the magnitude of the balance disturbance caused by the moving room is much higher in inexperienced walkers than experienced walkers. Newly walking infants are often literally ‘bowled over’ by the imposed global optic flow. The development of responsiveness to PLOF provides an example of the role of experience in functionalizing visual information for the control of posture. Utilizing the moving room paradigm, it was found that 5-month-old infants were essentially unresponsive to PLOF when exposed to side-wall movement in a moving room (Bertenthal and Bai, 1989). The same lack of responsiveness was reported in 7 month olds (Higgins et al., 1996). In contrast, both studies noted major changes in responsiveness between 7 and 9 months of age, changes that the authors speculated were related to experience with independent mobility. The link between mobility experience and responsiveness to PLOF was subsequently affirmed in a second experiment (Higgins et al., 1996). Eight-month-old infants who were proficient at crawling on hands and knees or moving about in wheeled walkers showed significantly higher postural responsiveness to side-wall motion in the moving room than same-aged infants who were not yet locomoting independently. These findings have recently been confirmed and extended using a randomized controlled trial (RCT) (Uchiyama et al., 2008). In the RCT, prelocomotor infants were taught to drive a baby go-cart (powered mobility device [PMD]) by pulling on a joystick mounted at the front of the device (Fig. 10.3). Infants were tested for responsiveness to PLOF before and after 15 days of training. A control group received the same pre- and post-testing, but no training. The findings were remarkably clear—the PMD infants’ responsiveness to side-wall motion in the moving room increased significantly as a function of their training, whereas the control group showed no changes in visual–postural coupling at all (Fig. 10.4). Equally importantly, the PMD-trained infants, as well as hands-and-knees crawlers and infants with ‘artificial’ walker experience who were tested in a preliminary study showed heightened emotional reactions to the imposed optic flow (Uchiyama et al., 2008). The latter findings are important because they suggest that the postural compensation induced by the imposed peripheral optic flow was more meaningful to the infants with locomotor experience than those without locomotor experience. Presumably, the meaningfulness came from the threat to stability that was associated with the imposition of the newly functionalized peripheral optic flow in the locomotor infants but not the prelocomotor infants. Figure 10.4 Changes in the coupling between side-wall movement in the moving room and postural sway before and after training in a powered mobility device in the Uchiyama et al. (2008) study. Another interesting finding was the increase in positive emotion (smiling) associated with learning to control the PMD (Uchiyama et al., 2008). The increase in smiling coincided with dramatic increases in the amount of pulling engaged in by the infant, implying that once the joystick had become functionalized as a means to an end (i.e., the infant understood that pulling the joystick caused the PMD to move forward), the infant had a heightened sense of agency—a heightened sense of control of the self relative to the environment. The consequences of PMD training for prelocomotor infants have been replicated and extended (Dahl et al., 2013). In addition to showing heightened responsiveness to PLOF, PMD-trained infants also displayed greater cardiac indications of wariness of heights when lowered to the deep side of a visual cliff (see Fig. 10.5). This finding is significant for researchers interested in the psychological consequences of motor activity because wariness of heights was one of the first psychological outcomes linked to experience with independent mobility (see Campos, 1976). In other words, the contemporary work on mobility and psychological development had its origins in the serendipitous and surprising discovery that infants did not display wariness of heights until they had acquired experience with independent locomotion. Figure 10.5 A child being lowered to the deep side of the visual cliff in the Dahl et al. (2013) study. Despite the clear role that locomotor experience has been shown to play in facilitating the use of peripheral optic flow for balance, there is evidence for a precocious competence in the neonate for responding to lamellar optic flow (e.g., Barbu-Roth et al., 2009; Jouen, 1988, 1990; Jouen et al., 2000). For example, Jouen et al. (2000) reported that neonates (3 days old), who were reclined in a specially designed infant seat between two television monitors, showed head retraction to a continuously moving optical pattern. This precocious sensitivity highlights a consistent finding in perceptual development—infants can detect information well before that information is useful for guiding action (Goldfield, 1995). Following Haith (1993), we have referred to perceptual, cognitive, and emotional biases and precocities as partial accomplishments. These biases are the rudimentary primitives from which functional skills are constructed. Consequently, locomotor experience appears to play a pivotal role in functionalizing these primitives for utilization in the control of skilled behaviour. Though visual proprioception will obviously improve as the visual system’s ability to pick up information improves via accommodation, convergence, decreasing thresholds for motion detection, oculomotor co-ordination, and widening of the field of view, locomotion presses the infant to differentiate global, radial, and lamellar optic flow. The press can be understood with respect to the demands placed on the visual system during locomotion. Gibson (1979) has noted that vision serves three important functions in mobility: (1) detecting that the surface supports locomotion; (2) steering through apertures and around obstacles; and (3) maintaining postural stability. Essentially, the individual must concurrently process information about self-motion and attend to the layout of surfaces and objects during locomotion. The newly locomoting infant is pressed to differentiate spatially delimited patterns of optical flow so that the various demands on the visual system can be accomplished successfully and efficiently (Gibson and Schmuckler, 1989). Relegating the control of postural stability to the peripheral field of view frees the central field of view to accomplish the task of surface scanning and steering and to focus on other features of the environment that might be important to secondary tasks that are nested into the primary task of locomotion: noting landmarks, for example. The overall process by which visual proprioception becomes more finely tuned involves the education of attention (Gibson, 1979) to specific patterns of information that are potentially usable for postural control, differentiation of spatially delimited regions of optic flow, and precise mapping, via self-produced experience, of specific patterns of optic flow onto specific neuromuscular control strategies. With experience, functionalization leads to increasing specificity in behaviour over time, which then increases the effectiveness, efficiency, and adaptability of behaviour to the task and to the environment in which it occurs. Because visual proprioception plays such an integral role in balance and because balance is so essential to the development of all skilled interaction with the environment, deficits in visual proprioception can seriously compromise children’s skill development. Moreover, given the contribution that visual proprioception makes to the development of a concept of the self and given how important self-awareness is to having a sense of agency, deficits in visual proprioception have serious implications for the child’s development of a sense of agency. Locomotion is more than simply organizing a pattern of movement to make forward progression. Functional locomotion requires that the individual has an intimate understanding of the spatial layout and his or her relation to it. In the previous section we noted that visual proprioception provides infants with one of the earliest means to differentiate themselves from the environment because specific patterns of optic flow are associated with their own movements versus movements of objects and surfaces. Improvements in visual proprioception refine the infant’s perception of orientation relative to the external surrounds and consequently feed into a deeper appreciation of external space. Optic flow will ultimately be used to gauge heading direction (Banton and Bertenthal, 1997; Gilmore and Rettke, 2003) and judge speed and distance after the onset of independent locomotion (Campos et al., 2000). However, functional locomotion also relies on navigation and memory. Maintaining an orientation to the external layout, knowing where we are going and where we have been, remembering landmarks, and recalling where objects are located or hidden once we have moved are all features of spatial cognition (Clearfield, 2004; McKenzie, 1987). The functional independence that comes with locomotion would be very limited indeed without spatial-cognitive skills. One of the most puzzling spatial-cognitive phenomena in infancy is the difficulty that infants have in searching correctly for an object hidden in one of two locations. Although infants between 8 and 9 months of age are able to successfully retrieve an object hidden within reach at one location, they often do not succeed when the object is hidden under one of two adjacent locations, even when the locations are perceptually distinct (Bremner, 1978; Piaget, 1954). More curiously, infants at this age will often fail to retrieve a hidden object after it has been moved to a new location, even though they have successfully retrieved it from the old location and they saw it being moved to the new location. Infants will surprisingly perseverate at searching in the old location. The perseverative search is referred to as the A-not-B error. The longer the delay imposed in searching for the object when it is hidden in a new place, the poorer the infant’s performance (Diamond, 1990). The A-not-B error gained widespread attention thanks to Piaget (1954), who used it to highlight the importance of action in the child’s intellectual development. Piaget (1954) believed that infants made the error, in part, because they coded the positions of objects egocentrically (relative to their own body) rather than allocentrically (relative to each other or to stable environmental features). Contemporary explanations suggest that many factors play a role in the error (Munakata, 1998; Smith et al., 1999). Regardless of the explanation, it is now clear that locomotor experience is related to the infant’s overcoming of the error (see Smith et al., 1999). A number of researchers, including Piaget (1954), have speculated about a link between skill in spatial search and locomotor experience (Acredolo, 1978, 1985; Bremner, 1985; Bremner and Bryant, 1977; Campos et al., 1978). However, Horobin and Acredolo (1986) were the first to test the link. Their results showed clearly that 31–41-week-old infants with greater amounts of locomotor experience were more likely to search successfully at the B location on a series of progressively challenging hiding tasks. These findings were replicated and extended using a similarly challenging series of spatial search tasks (Kermoian and Campos, 1988). The tasks ranged from retrieving an object partially hidden under a single location to the A-not-B task with a seven-second delay between hiding and search. The infants in the study were all 8.5 months of age, but differed in the amount of experience each had with independent mobility; one group was prelocomotor, a second group was prelocomotor but had experience moving in an infant walker, and a third group had experience with hands-and-knees crawling. The results showed clearly that infants with hands-and-knees crawling experience or walker experience significantly outperformed prelocomotor infants on the spatial search tasks. Moreover, search performance improved with greater amounts of locomotor experience. For example, 76% of hands-and-knees crawling and walker infants with nine or more weeks of locomotor experience successfully searched in the B location on the A-not-B test with a three-second delay compared to only 13% of prelocomotor infants. These findings have been replicated and extended (Tao and Dong, 1997), using cross-sectional and longitudinal investigations, and taking advantage of ecologically and culturally mediated delays in the onset of independent mobility in urban Chinese infants. Specifically, infants in Beijing who were delayed in locomotion by two to four months relative to North American norms initially performed poorly on the A-not-B test, then improved dramatically as a function of locomotor experience, regardless of the age of onset of locomotion. Even more pertinent to the current chapter, aspects of these findings have been confirmed in a longitudinal study of seven infants with spina bifida who were delayed in the onset of locomotion relative to typically developing infants (Campos et al., 2009). The infants were tested on a two-position hiding task in which a toy was hidden only in one location, with a second hiding location serving as a distractor. The infant’s ability to follow the gaze and point of an experimenter toward an object that was outside their field of view was also tested. Note that an ability to follow the point and gaze, also referred to as referential gestural communication, has also been linked to experience with locomotion. The infants were tested monthly after they were recruited into the study until two months after the delayed onset of independent locomotion, which occurred at 10.5 months of age in four of the infants and 8.5, 11.5, and 13.5 months of age in the other three infants. The results revealed dramatic improvement on the task following the onset of locomotion. Infants searched successfully for the hidden object on only 14% of trials while they were prelocomotor, but improved to 64% correct search following delayed onset of locomotion (Fig. 10.6). Similarly, in the referential gestural communication task, the infants were much more likely to look in the correct direction of the experimenter’s gaze and point and less likely to look at the experimenter, following the delayed onset of locomotion. Figure 10.6 Changes in spatial search performance following the delayed onset of locomotion in infants with spina bifida in the Campos et al. (2009) study.
The consequences of independent locomotion for brain and psychological development
Brief historical perspective on motor activity and psychological functioning
The neglect of motor activity’s role in psychological functioning
A resurgence of interest in motor contributions to mind
Independent mobility and psychological development in the human infant
Visual proprioception: what is it and why is it important?
The development of visual proprioception

Responsiveness to global optic flow
Responsiveness to peripheral lamellar optic flow (PLOF)

How does mobility experience contribute to changes in visual proprioception?
The development of spatial understanding
Manual search for hidden objects
Related posts:
 B: Findings from a series of studies of passive mechanical properties of muscle in young children with cerebral palsy
B: Findings from a series of studies of passive mechanical properties of muscle in young children with cerebral palsy
 C: Kick start: using the mobile infant learning paradigm to promote early leg action
C: Kick start: using the mobile infant learning paradigm to promote early leg action
 Treadmill training in early infancy: sensory and motor effects
Treadmill training in early infancy: sensory and motor effects
 Very early upper limb interventions for infants with asymmetric brain lesions
Very early upper limb interventions for infants with asymmetric brain lesions
![]() Corticospinal tract development and activity-dependent plasticity
Corticospinal tract development and activity-dependent plasticity
 Training lower limb performance in early infancy: Support, balance and propulsion
Training lower limb performance in early infancy: Support, balance and propulsion
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


The consequences of independent locomotion for brain and psychological development

