INTRODUCTION
Therapeutic exercise encompasses basic physiology and physiologic sciences as well as exercise science. This chapter briefly outlines the use of physical activity, muscular contraction, and physical exertion in order to prevent, treat, and rehabilitate physical conditions as well as improve performance or fitness. Exercise can be used to increase or improve strength, endurance, aerobic capacity and conditioning, flexibility, body mechanics, and proprioception, and can help a patient progress toward functional movements and activities. Although the chapter summarizes essential knowledge, readers are encouraged to explore additional sources for more comprehensive details.
AEROBIC & ANAEROBIC METABOLISM
Adenosine triphosphate (ATP) is the primary energy substrate used by muscle cells (myocytes) to generate contractile force during exercise. When the amount of intramuscular ATP stores is depleted after 10 seconds of exercise, it must be replenished. ATP regeneration occurs through three processes, using aerobic (ie, inclusive of O2) and anaerobic (exclusive of O2) avenues. All three ATP generation systems occur simultaneously. Activities that are shorter in duration and have greater intensity rely on anaerobic systems, whereas those of lower intensity and longer duration rely on aerobic systems.
The two anaerobic systems fueling muscle activity are the adenosine triphosphate–phosphocreatine (ATP-PC) system and anaerobic glycolysis. Of the two, the ATP-PC system regenerates ATP more quickly because it involves only a single chemical reaction. In this reaction, the bond between phosphate (P) and creatine (C) in phosphocreatine (PC) is broken, releasing energy and a phosphate group to resynthesize ATP from ADP: PC + ADP → ATP + P. Unfortunately, the PC stores of myocytes can sustain only about 30 seconds of activity. Activities involving short, powerful bursts (eg, the swing of a baseball bat, a 100-m sprint, or powerlifting) primarily rely on the ATP-PC system.
Glycolysis, the second aerobic system, involves a series of reactions whereby simple (glucose) and long-chain (glycogen) carbohydrates are metabolized into pyruvate. During both aerobic and anaerobic glycolysis, ATP and pyruvate are synthesized. However, during anaerobic conditions, pyruvate is preferentially fermented into lactate so that glycolysis, and its ATP generation, can continue. Anaerobic glycolysis is, nevertheless, limited in its ability to generate ATP because the acidification of cytosol by lactate eventually impedes glycolytic enzyme activity. Interestingly, although lactate can impede glycolysis, some lactate is transferred to the systemic circulation. There, lactate may be converted to glycogen in the liver through the Cori cycle. Alternatively, intracellular and extracellular lactate may be oxidized into pyruvate and then metabolized for ATP in a process known as the lactate shuttle. Anaerobic glycolysis produces more ATP over a longer time frame than the ATP-PC system; however, both anaerobic systems generate less than 10% of the body’s potential ATP. Highly intense, short-duration exercises of up to 2 minutes, such as 400-m sprints or repetitive weight lifting, will exhaust both systems.
On the other hand, in aerobic conditions, pyruvate is metabolized into acetyl-coenzyme A (CoA) before entering the tricarboxylic acid (TCA) cycle in the mitochondria. Noncarbohydrate energy sources, such as fats and proteins, are also catabolized into the acetyl-CoA substrate, but at slower rates than carbohydrates. The TCA cycle produces ATP as well as most of the energy-rich oxidizing agents used in oxidative phosphorylation through the electron transport chain. The remaining 90% of ATP is generated by means of oxidative phosphorylation, which is the chief source of ATP production after exhausting the ATP-PC and anaerobic glycolytic systems. This occurs in endurance activities lasting more than 3–4 minutes, such as long-distance running over a distance of more than 1 mile and cross-country skiing events.
MUSCLE STRUCTURE, PHYSIOLOGY, & BIOMECHANICAL PRINCIPLES
The potential energy of ATP must be converted into mechanical force to create movement during exercise. Such a process occurs in muscular tissue with great efficiency. Muscles are composed of fascicles containing myocytes. Contraction of the myocytes relies on the extensive network of myofibrils filling the cytoplasm. A myofibril is either a thick myosin myofilament, or a thin actin myofilament. Both types of myofilament overlap near each other in units called sarcomeres, as shown in Figure 9–1.
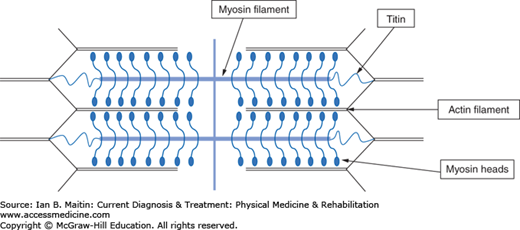
The sarcomeres line up end to end (which, in skeletal muscle, accounts for its striated appearance). Muscles contract by means of an overlapping phenomenon involving sliding filaments. For contraction to occur, calcium ions must first be released into the cytoplasm from their storage location in the sarcoplasmic reticulum of the myocyte. Actin is composed of troponin and tropomyosin molecules; in the absence of calcium, tropomyosin blocks the myosin head from interacting with actin. However, when calcium is present it binds to troponin, preventing tropomyosin from interfering with the myosin binding site on actin. This frees troponin to bind myosin. ATP can then be converted to energy and ADP at the myosin head. The newly released energy breaks the bond at the actin–myosin head, allowing the myosin head to bind to another actin site, contracting the sarcomere length, and thus the myocyte and muscle tissue length. Force is generated when the muscle contracts against load.
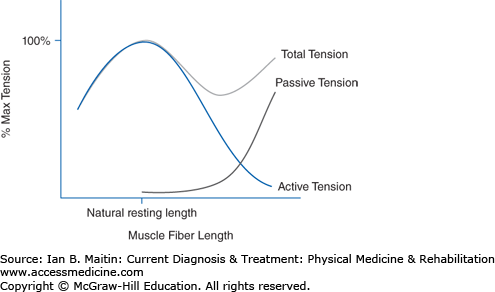
The optimal muscle length for active force generation is known as the natural resting length. At the level of the sarcomere, this is the length at which actin and myosin overlap maximally without collision of opposing actin filaments, resisting further contraction. A muscle is typically near natural resting length at the midrange of joint motion if the muscle acts on that joint. The total tension generated by a muscle is the sum of active muscle tension and passive elastic tension. Active tension is created by actin–myosin head interaction; passive tension is derived mainly from titin fibers that connect the myosin to actin filaments, as seen in Figure 9–2. At lengths shorter than neutral resting length, total tension is simply a factor of active contraction of the muscle. But as the muscle is lengthened, the total tension is the combination of both active and passive tension. Active tension decreases at lengths greater than neutral resting length, whereas passive tension continues to increase. In general, the maximum overall tension within a muscle is at a length slightly longer than its resting length.
Skeletal muscle fibers do not all contract in the same manner, and researchers have divided fibers into subtypes based on contraction speed and endurance. Each subtype varies in how quickly ATP is consumed, sources of fuel utilized for ATP, speed of muscle contraction, and rate of fatigue. The two categories are type I (slow twitch) and type II (fast twitch). Type I fibers generally have fewer myofibrils in their cytoplasm, which results in less muscle bulk and force generation. Myosin head–actin crosslinking and ATP usage is also two to three times slower in type I fibers compared with type II. However, type I myosin fiber contractions can last up to ten times longer than those of type II. Type I fibers possess significant amounts of mitochondria, oxygen-binding myoglobin, and capillary access. These factors give the muscle fibers a distinctive dark-red color and, more importantly, mean that they excel at oxidative phosphorylation. Type II fibers, which lack the mitochondria and myoglobin content of type I fibers, rely more heavily on ATP-PC and anaerobic glycolysis for fast ATP production. These fibers have copious actin–myosin filaments, creating greater cross-sectional muscle fiber volume, and also force production. It is apt to state that type II fibers are designed for sprinting, whereas type I are geared for endurance activities.
Although this binary distinction between fiber types is useful conceptually, in reality a continuum of fibers shares characteristics of both fiber types. Additionally, type II fibers are subdivided into three categories, designated IIa, IIx, and IIb. Type IIa fibers, which produce greater force than type I, are relatively fatigue resistant and have high oxidative and glycolytic capacities. Type IIx fibers produce greater force at the cost of slightly less oxidative capacity and fatigue resistance. Finally, type IIb fibers are most similar to the type II fibers discussed earlier, with high glycolytic and force generation abilities but poor oxidative capacity and fatigue resistance.
A concentric contraction is a muscle-shortening contraction. When the force generated by the muscle is sufficient to overcome resistance during the contraction, the muscle will shorten. An eccentric contraction is a muscle-lengthening contraction. This occurs when the force generated by the muscle is insufficient to overcome the external load and the muscle lengthens despite contraction. This type of contraction can also be used to decelerate and control movements of limbs and joints as well as to control the movement of external objects being lifted or carried. An isometric contraction is a contraction of the muscle that involves neither muscle shortening nor lengthening (ie, no joint movement). The tension in the muscle equals the load; thus, no movement is created. The length of the muscle during contraction has an impact on the force it can generate during isometric contraction. The natural resting length of the muscle (described earlier) yields the highest isometric force, and when the length from this neutral point either increases or decreases, the isometric force will decrease.
The muscle–tendon insertion site, angle of the insertion, and distance of insertion from the center of joint rotation all affect muscle leverage, which can be represented by torque. In biomechanics, torque is the product of the distance of a force from the center of joint rotation and the magnitude of the perpendicular force vector. If the insertion angle of the tendon is at 90 degrees (ie, perpendicular to the bone), then torque production is the greatest for a given tension in the muscle. As the angle increases or decreases from 90 degrees, the torque will also decrease. Tendon insertion distance from the center of rotation also affects torque. Insertion distances further from the center of rotation increase the torque produced by a muscle for a given contractile force, but at the expense of total excursion of movement, or angular range. For a given contraction, the muscle will shorten a certain amount, and if tendon insertion is further from the center of rotation of the joint, the excursion of movement of that joint will be less than if the tendon inserted closer to the center of rotation. Slight anatomic differences in muscle–tendon or fiber insertion points may allow different force or torque production for a given contractile effort and could account for variability in performance capabilities.
Variability in muscle fiber alignment also affects the force generation and contraction speed of a muscle. Alignment is defined as the angle between the muscle fibers and a line drawn between the origin and insertion of the entire muscle, which is typically the plane of force generation. The simplest arrangement exists when fibers run parallel to the force generation plane; these are called fusiform fibers. Compare this with pennate fibers, in which there is a 5- to 30-degree angle between the fibers and the force generation vector. This arrangement gives pennate fibers an advantage over fusiform as it allows for higher densities of fibers in a cross-sectional area. Comparing cars parked in parallel with cars parked diagonally provides an analogy; more cars fit within a given space when parked diagonally rather than parallel parked. With muscles, however, greater force generation comes at the cost of overall muscle contraction speed, as each fiber needs to contract a longer distance to shorten the entire muscle. Although fusiform fibers may have a smaller cross-sectional fiber area, they are capable of contracting significantly faster than pennate fibers. Examples of pennate muscles include the soleus and vastus medialis muscles. Sartorius and biceps brachii muscles have a fusiform arrangement.
Contraction velocity also affects torque. In the early 20th century, Hill proposed a force–velocity relationship showing that muscle torque is greatest during isometric contraction and decreases hyperbolically as contraction velocity increases. This theoretical curve is represented in Figure 9–3, with force expressed as torque.
Figure 9–3
Hill’s proposed force–velocity curve. Eccentric contractions have only shown greater force production compared with isometric contraction in laboratory modeling, as predicted by Hill (dotted line). However, physiologically, maximal eccentric force production is achieved at approximately isometric levels (solid line).
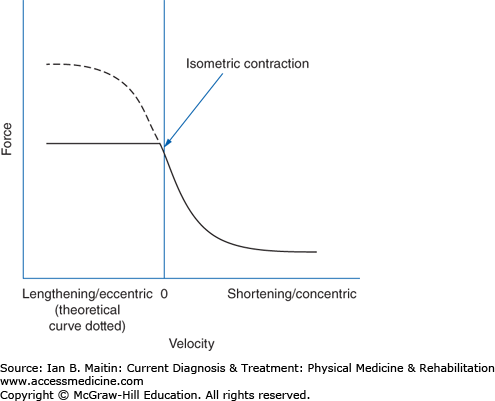
The isometric and concentric portion of this model has been supported experimentally; for example, isokinetic devices have measured muscle-generated torque showing that there is less torque as the velocity of concentric contraction (measured in revolutions per minute) increases. Eccentric contractions have, at least in vivo, been shown to produce significantly greater forces than isometric contractions. In situ, voluntary eccentric muscle contraction force in humans supersedes isometric contraction force only minimally, if at all, and may be limited by a neurologic tension-regulating mechanism to protect the muscle tissue. Additionally, despite diminished contractile force at higher concentric contraction velocities, muscular power (force times velocity) increases.
NEUROMUSCULAR PRINCIPLES
A motor unit consists of a group of myocytes and the α motor neuron axon innervating them. The muscle fibers within a motor unit contract simultaneously when an action potential propagates from the α motor neuron’s cell body and down its axon, causing acetylcholine release from the presynaptic axon terminal into the neuromuscular junction. Acetylcholine is a neurotransmitter that binds the postsynaptic nicotinic acetylcholine receptors on the postsynaptic myocyte membrane surface, inducing calcium release from the sarcoplasmic reticulum of the postsynaptic myocyte. The end result is a contraction, as described previously by the sliding filament theory. Different motor units are dispersed throughout the cross-sectional area of a muscle. Each motor unit varies in numbers of fibers and fiber subtype composition. The overall magnitude of a muscle contraction is thus not necessarily proportional to muscle bulk, but rather reflects the number, size, and fiber composition of the motor units that are contracting simultaneously. The number of fibers per motor unit is variable and dependent on the size and function of the muscle. Muscles that perform fine control and small-coordinated movements, such as the lumbricals, contain only a few fibers per motor unit. This is unlike muscles that perform gross movements and activity, such as the gastrocnemius, which have many fibers per motor unit.
The central nervous system asserts precise control over how many and what types of motor units are activated during bulk muscle contraction. Not every muscle fiber is activated at once. Histologically, fast-conducting motor neurons innervate motor units with a greater percentage of type II fibers while slower, smaller motor neurons innervate motor units with more type I fibers. Henneman’s size principle states that motor units containing the fewest myocytes and most type I fibers are recruited (or activated via action potential) first for any muscle contraction. As demands and the need for higher force production arise, larger motor units with increasing amounts of type II fibers are recruited. The benefit of recruiting the smaller type I dense motor units first is that a contraction can be sustained for the longest amount of time by relying on oxidative phosphorylation energy production rather than anaerobic processes. Further recruitment of motor units with stronger contractility also enables fine control over the amount of force generated. In addition to gradually selecting motor units with larger sizes to generate force, motor unit action potentials also become more frequent, and contraction frequency increases.
If motor unit recruitment is maximized, the muscle is said to display its maximum voluntary contraction (MVC). Although a muscle may be able to activate all motor units of the muscle at once (a point that is debated) to achieve a MVC, the maximal force produced by the muscle is intimately related to the muscle’s cross-sectional area. The absolute muscle strength per cross-sectional area varies somewhat, depending on the source consulted, but historical values average around 15 N/cm2.
Muscle fatigue can be defined generally as any reduction in the maximal force-generating capacity. It is an active impairment of motor performance manifested by either an increased sense of effort to maintain desired force output, or an overall involuntary decline in force output. Several factors other than muscle fiber type composition lead to muscle fatigue.
Inadequate central drive involves a decrease in motor unit recruitment for MVC following a fatiguing task. Muscle fiber membrane excitability decreases with continuous stimulation, thus creating a failure of action potential propagation. Contractile force may decline without a change in EMG activity, an occurrence termed excitation–contraction coupling failure. With sustained muscle contraction, metabolic byproducts can accumulate locally, resulting in intramuscular acidosis. This affects the myofibril cross-bridges, leading to a decline in the number of cross-bridges that can be formed as well as the force exerted by each cross-bridge. At exercise levels below maximum aerobic intensity, but for long durations (> 1 hour), this process will lead to glycogen depletion within the muscle.
Muscle fatigue is reversible, but recovery time is variable depending on the cause of that fatigue. Short-duration, intermittent, submaximal contractions may result in fatigue related to excitation–contraction coupling failure, with a slow recovery of up to 30–60 minutes. In contrast, fatigue from the accumulation of metabolites appears to occur rapidly during maximal or near-maximal contractions but has a recovery time of less than 2 minutes.
Finally, the individual’s current fitness level and psychological motivation level also affect the rate of development of fatigue. Training can reduce the fatigability of a muscle, but the response seems to be task specific. Of note, since muscle endurance training appears to be task specific, clinical endurance tests using measures such as an isokinetic device may have little carryover to functional activities.
PRINCIPLES OF AEROBIC CONDITIONING
Vo2 is defined as the total amount of oxygen consumption from arterial blood per unit time. It can be represented by the Fick equation, in which the difference between arterial and venous oxygen content is multiplied by cardiac output:
As aerobic intensity increases, more arterial oxygen is used for oxidative phosphorylation, and Vo2 increases. This continues until a maximum is reached, in which there is no further increase in oxygen uptake despite increases in exercise intensity—the Vo2max. This value represents the maximal amount of oxygen consumption from the blood per unit time and is thought to be the most accurate measure of an individual’s aerobic capacity for a specific activity. Vo2max is affected by many variables, including type of exercise, effort of the individual, training status of the individual, predominant muscle fiber type used in exercise, and adaptations to aerobic exercise, such as capillary vascularization within muscle.
A metabolic equivalent (MET) is a unit that is used to estimate, generally, the metabolic cost of a physical activity. It is depicted as a multiple of the resting metabolic rate, and thus is a multiple of the Vo2 found at rest. One MET equates to approximately 3.5 mL of O2/kg/min, or 250 mL/min in males and 200 mL/min in females of average sizes. For greater accuracy in individuals of varying mass, the MET may also be expressed as Vo2 divided by body mass: mL/(min × kg). METs are often used in place of Vo2 to easily gauge the relative intensity of a given physical activity.
Both aerobic and anaerobic conditioning rely on the overload principle, which states that increasing frequency, duration, or intensity when performing an activity (mode) will stimulate physiologic adaptations furthering an individual’s capacity to perform the same activity. A progressive training program should be structured to increase at least one of these components over time to produce exercise gains.
The specificity principle refers to the specific adaptations that are made to imposed demands on the system (SAID principle), which occurs in both neurologic and biomechanical overload. Strength and endurance gains made during overload training with a specific exercise modality are realized in activities most similar to that modality. For example, optimal training for an activity should include the same muscle groups, joint ranges of motion and positioning, aerobic intensity, and duration as are used in the activity. This has been referred to as functional training, in which motions and actions in training replicate those that will be performed during the sport or competition.
All aerobic physiologic gains made through training are reversible and can be quickly lost as a result of detraining if the individual ceases to perform the specified activity. Losses in Vo2max and muscular capillary density have been reported in trained individuals confined to bed rest for several weeks, and heart size decreases to pretraining levels when endurance training ceases.
AEROBIC TRAINING
Related posts:




Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


