17 The Thorax The heart is roughly the size of a fist and weighs approximately 300 g. It is divided by the cardiac septum into a right and a left half, which usually have no connection with each other. In each half, we find an atrium and a ventricle, separated from each other by a flap-shaped valve (atrioventricular or AV valve). A different type of valve is found in the vessels that lead out of the ventricles—the pocket valves (semilunar valves). Both valves are infoldings of the inner layer of the heart, the endocardium. They function as valves by allowing the inflow of blood in one direction but preventing a backflow in the opposite direction. The oxygen-depleted blood from the body’s circulation flows through the superior and inferior vena cavae into the right atrium. The superior vena cava (SVC) is a central vessel that collects the blood from the upper extremities and the areas of the head and neck. The inferior vena cava (IVC) delivers blood from the abdomen, the lesser pelvis, and the lower extremities. Through the tricuspid valve, the flap valve of the right heart (three flaps), the blood flows into the right ventricle and from there on through the pulmonary valve into the pulmonary artery. In the capillary net of the lungs, the deoxygenated blood is rearterialized and then flows through the pulmonary veins to the left atrium. After passing through the mitral valve (two flaps), the blood flows into the left ventricle and then on through the aortic valve into the aorta and the rest of the body. The heart functions as a combined pressure-and-suction pump in a closed system. The wall of the heart consists of three layers. On the outside is the epicardium, a layer of connective tissue that is followed by the pericardium. The middle layer consists of a meshwork of interconnected striated muscles (syncytium). This special structure guarantees that any impulse in the heart is transmitted from cell to cell. The muscles of the left ventricle are roughly three times as thick as those of the right ventricle. The reason for this is the difference in pressure that must be built up to expel the blood from the ventricles: the pulmonary circulation is characterized by a pressure of approximately 25 mmHg; the systemic circulation has to overcome a systolic pressure of approximately 120mmHg. The innermost layer of the heart is the endocardium, which is also formed of connective tissue. The cardiac impulse-conducting system consists of specially built cardiac myocytes that have the ability to autonomously create impulses. The heart’s pacemaker is the sinoatrial node, a roughly bean-sized knot at the entrance of the SVC in the right atrium. It generates impulses with a frequency of 70–80/min. After a diffuse transmission through the atria, the AV node receives the impulse. This structure is located at the border between the atrium and ventricle in the right heart. From there, the action potential is conducted to the bundle of His on the right side of the interventricular septum. Then there is a division into two ventricular branches that continue onto the muscles of the right and left ventricles and end in the Purkinje fibers. The heart is located in the mediastinum inside the pericardium, with the tip pointing toward the front at the bottom left and the base toward the back at the top right. The right half of the heart faces the anterior thoracic wall, the left points posteriorly. The right border of the heart projects at the sternal attachment of ribs 2–6—approximately 2–3cm away from the sternum—onto the anterior thoracic wall. The caudal border is formed by the diaphragm. The left border of the heart runs as follows: cranial for a distance of approximately 2cm from the sternal attachment of the second rib in a diagonal course to the tip of the heart in the fifth intercostal space approximately 2cm medial to the medioclavicular line. Posteriorly, the most caudal point of the heart lies at the level of T10 during exhalation and shifts downward by a distance of one and a half vertebrae during inhalation. Lateral Anterior Posterior Caudal. Diaphragm. Via the pericardium, the heart is firmly attached in the mediastinum. Due to its structure as a serous cavity, the pericardium also ensures frictionless and free moveability during heart activity. Cranially, the heart is suspended by the entering and exiting vessels. The pericardium has multiple attachments in all directions: There are connections to the esophagus, bronchial tubes, and pulmonary veins. Laterally, the pericardium is linked via connective tissue to the parietal pleura. The pericardium and its attachments are incorporated into the fascial system of the “central tendon,” which extends from the base of the skull to the lesser pelvis. Fig. 17.1 Location of the thoracic organs. A right and a left coronary artery originate at the aortic sinus (dilatation at the origin of the aorta above the aortic valve). During diastole, the pockets of the aortic valve fill up, and blood fills the coronary arteries. In functional terms, the coronary arteries are endarteries, i.e., they do not form anastomoses with each other: if a branch of the artery is occluded, the myocardial tissue supplied by this vessel perishes. The lymph flows to the anterior mediastinal lymph nodes in front of the tracheal bifurcation and to the lymph nodes in the vicinity of the large vessels by the heart. Fig. 17.2 Ligamentous structures of the pericardium. Both vegetative sections meet in the cardiac plexus. The plexus surrounds the aorta and the roots of the other large vessels in the vicinity of the heart; from there, vegetative nerves run together with the coronary arteries. Maximal time: 11 a.m.–1p.m. Minimal time: 11 p. m.–1 a.m. For basic information, see page 34. The two halves of the lung largely fill the lateral chest. The left half of the lung is slightly smaller because the heart claims some of the space in the left chest. The left half of the lung consists of two lobes that are divided into nine segments. The right half consists of three lobes and ten segments. Each segment is supplied with air by a segmental bronchus. These bronchial tubes combine to form the lobar bronchi, which then form the main bronchus. The two main bronchi together form the trachea. The actual location of gas exchange is in the alveolae. The human body has approximately 300–400 million alveolae with a total surface area of 100–140 m2. The lung is surrounded by the visceral pleura, which runs into the parietal pleura in the suspension area of the lung. The fluid-filled space between the lung and pleura is highly significant for the frictionless mobility and fixation of the lung. The cavity between the two pleural layers serves partly as extra space into which the lungs can expand during deep inhalations. The borders of the pleura are therefore listed separately from the borders of the lungs. Fig. 17.3 Topography of the two halves of the lung and pleura: anterior view. Fig. 17.4 Topography of the two halves of the lung and pleura: posterior view. Cranial Approximately 3 cm above the first rib, tough and thickened. Anterior and Medial Behind the sternum, on the left with cardiac impression. Posterior and Medial T1–T12, paravertebral. Caudal Medioclavicular line: rib 7 Anterior axillary line: rib 8 Axillary line: rib 9 Posterior axillary line: rib 10 Scapular line: rib 11 Paravertebral: rib 12 The cranial and paravertebral borders coincide with those of the pleura. Caudally, the lung sits one or two intercostal spaces above the borders of the pleura during average inhalations. In deeper in- or exhalations, the lung moves downward or upward by about one intercostal space. On the whole, the left lung sits slightly lower because the liver displaces the lung cranially on the right. Left Oblique Fissure This starts in the back near the fourth costovertebral joint and, after a diagonal course around the chest, ends anteriorly in the vicinity of the sixth sternochondral joint. Anteriorly it is closely connected to ribs 5 and 6, posteriorly to ribs 4 and 5. Right Oblique Fissure This starts posteriorly near the third costovertebral joint and ends anteriorly in the vicinity of rib 6. Anteriorly, it is closely connected to rib 6, posteriorly to ribs 3–6. Horizontal Fissure of the Right Lung This originates posteriorly in the oblique fissure below the scapula at the level of rib 4/5 and ends anteriorly slightly below the third sternochondral joint. This joint and rib 4 are located in close proximity to the fissure. The adhesive force caused by the vacuum in the pleural cavity ensures that the lung does not contract toward the hilum as the result of its own elasticity. The parietal pleura is firmly attached to the inside of the chest: fascial attachments to the sternum and ribs are found along its entire course. The pleura is also attached solidly to the diaphragm. Toward the mediastinum, it covers the neighboring organs without establishing a firm tie. At the hilum of the lung caudally, the pulmonary ligament is formed as a fold of the parietal pleura. This ligament is attached to the diaphragm. At the pleural dome, we find suspensory ligaments that connect the pleura to the first rib and the vertebrae C6–T1: The pulmonary artery and vein, the vasa publica (i.e., the vessels in charge of gas exchange), branch out from the bronchial tree to reach the capillaries that form a mesh around the alveoli. The vasa privata (i.e., the vessels that supply the lung tissue itself with oxygen and nutrients) originate as follows. Bronchial branches Bronchial veins. They run into the pulmonary veins or into the azygos and hemiazygos veins. Sympathetic Nervous System from T1–T2 to T5–T6 The fibers run to the pulmonary plexus and then on along the bronchial tree into the periphery of the lung. Parasympathetic Nervous System Vagus nerve. Maximal time: 3–5 a.m. Minimal time: 3–5p.m. For basic information, see page 34. The term “mediastinum” refers to the space in the chest that is bordered as follows. Anterior. Sternum. Posterior. Spinal column. Cranial. Upper thoracic aperture. Caudal. Diaphragm. Fig. 17.5 Topography of the mediastinum: sagittal view. Lateral. Both halves of the lung. In this space, we find a large number of important structures that are essential for the vitality of the entire body: These organs and circulatory structures are linked to each other by connective tissue. This ensures good fixation in the mediastinum. However, sufficient mobility must be present to follow the movements of the torso, arm, and head and neck, e. g., the esophagus and other organs must be able to stretch in a craniocaudal direction during a neck extension. Another factor that requires mobility is the expansion of the lung and the movements caused by diaphragmatic breathing. The mediastinum thus experiences alternating pushing and pulling. Lastly, heartbeats, in the sense of oscillations, also have an impact on the mediastinal structures. Thus we can see that continuous, even if partly only minor, movements in this apparently motionless space affect the organs of the mediastinum. This fact is particularly significant for the blood flow back into the heart, which is influenced by the suction effect of respiration, and for the nerve structures that are stimulated in the osteopathic sense by this constant movement. The mediastinum is tied into the fascial system of the “central tendon.” It constitutes the thoracic aspect of a fascial pull that reaches from the base of the skull down to the lesser pelvis. As a result, we can see fascial structural adaptations in the mediastinum that could lead to symptoms in the thorax but have their cause in a different location in the body. As a result of the vital importance of the mediastinal structures, abnormal fascial pulls can lead to significant functional changes. Here, we might consider the vagus innervation or the clinical picture of a hiatus hernia. Fig. 17.6 Topography of the mediastinum: transverse view. Respiration is the motor for the regular movement of the thorax. An average of 12–14 breaths/min require the chest to expand and contract rhythmically in its sagittal and transverse diameters. In biomechanical terms, we distinguish between two movement directions of the ribs: the rotational axis of the upper ribs that runs through the costotransverse and costovertebral joints lies almost parallel to the frontal plane—during inhalation, the result is mainly an expansion of the sagittal diameter of the chest. The rotational axis of the lower ribs lies almost in the sagittal plane. By raising the ribs during inhalation, the result is thus primarily an enlargement of the transverse diameter of the thorax. The central ribs have a movement axis that forms a 45° angle to the sagittal plane. Inhalations here lead to an expansion in the sagittal and transverse diameters. In the sternum, the movement of the ribs causes a rise cranially and an increase in the distance to the spinal column—the sternum moves anteriorly and superiorly during inhalation. Movements therefore occur in both the sternocostal and the chondrocostal joints. In the chondrocostal junction, the rib cartilages experience a torsion that is of great significance for the elastic and passive return of the thorax from the inhalatory position to the respiratory rest position. Inhalation is a process that is directed by respiratory muscles: easy respiration involves the diaphragm and the scalene and intercartilaginous muscles. These extend—as described above—the chest in its sagittal and transverse diameters; the diaphragm increases the thorax diameter caudally and raises the lower ribs. The contraction of the diaphragm causes a movement caudally while pushing the abdominal organs inferiorly and anteriorly. The movement anteriorly results from the soft abdominal wall, which does not provide active resistance against the displacement of the abdominal organs during inhalation. In easy respiration, exhalation is a passive process, directed by the elastic restorative force of the thorax. In deep inhalations, additional muscles assist in the expansion of the thorax. These accessory inhalatory muscles include the: Deep inhalations cause an extension in the spinal column, as a result of which the extensors of the spinal column can also be included indirectly among the inspiratory muscles. Forced exhalations likewise involve further accessory expirators: Here, we describe the mechanical heart action of the left heart. The same processes take place in the right heart. As a result of the contraction of the ventricle, intraventricular pressure rises. When this pressure is greater than the pressure in the atrium, the AV valves close (the semilunar valves are still closed). Tendinous fibers and papillary muscles prevent the AV valves from blowing through into the atrium. The surfaces of the individual flaps are greater than the opening to be closed. By broadly juxtaposing the edges of the flaps, closure of the valve is ensured even when the ventricular size changes. In the ventricle, no change in volume occurs, but only a reshaping of the ventricle into the form of a ball (= isovolumetric contraction). All muscle fibers change their length actively or passively. Duration of this phase: 60 ms when the body is at rest. This phase starts when the pressure in the left ventricle is greater than the diastolic pressure in the aorta (80mmHg). The semilunar valves open and pressure continues to rise until it reaches the systolic blood pressure value (approximately 120–130 mmHg). Finally, the ventricular contraction is released and the pressure drops back down. When the pressure is lower than the aortic pressure, the semilunar valve closes and systole is thereby concluded. During rest, approximately half the contents of the ventricle (130 mL) is ejected (= stroke volume). For approximately 50ms, all valves are closed. Ventricular pressure drops to almost 0 mmHg—when it falls below atrial pressure (2–4 mmHg), the AV valves open. The ventricle fills up in a quick passive phase (see below), caused by the atrial contraction taking place in this phase. With normal heart frequency, the filling of the ventricles is almost completed by the time of the atrial contraction—it receives 8% additional filling by it. The rapid fill phase of the heart begins during the ejection phase. As we have seen above, blood is pressed out of the ventricle. At the same time, blood is also sucked into the atrium. The reason for this is that, via the pericardium, the heart is firmly attached to the diaphragm—the apex of the heart is a punctum fixum. The atria are fixed by the vessels and hence also serve as a punctum fixum. The level of the valves is the punctum mobile. Ventricular contraction moves the heart valves in the direction of the apex. The slackened atria are stretched, which creates suction on the supply vessels of the atria; they fill with blood. As the ventricular muscles slacken, the valves return to their starting position and the AV valves open. As an inert mass, the atrial blood remains in the same place and the ventricle slides on top of this blood; the rapid passive filling of the ventricle results. This mechanism becomes especially important in heightened frequency with shortened diastole. First Sound = Sound of Contraction Second Sound = Valve Sound Third Sound Sudden jerk of the ventricular wall caused by blood rushing in during the fill phase of the heart—audible in children. Fourth Sound Contraction of the atria—at the end of the P wave in the ECG; physiological in youth, but in adults a sign of increased atrial strain. The venous backflow into the heart is not constant. When lying down, for example, it is greater than when standing up, due to the effects of gravity. Blood pressure varies almost by the minute. The heart must be able to respond to these constant changes in inflow in such a way that a steady bloodstream without blockages is guaranteed. The two potential disturbances in the continuous blood flow are pressure overload and volume overload. For example, infusion or valvular insufficiency. An increase in end-diastolic filling of the heart causes a greater stroke volume due to self-regulation. This is known as the Frank–Starling law of the heart. For example, rise in blood pressure or valvular stenosis. The ventricle has to work against increased pressure, and the adaptation occurs in steps: In chronic pressure or volume overload, the heart hypertrophies up to approximately 500 g. As a result of the end-arterial supply of the heart, cardiac insufficiency is a threat. The heart operates exclusively under aerobic conditions; similar to the skeletal muscle, it is unable to work without oxygen. During rest, it consumes approximately 10% of the total oxygen consumption in the body and during physical activity up to 40 %. The heart uses up 75 % of the oxygen in the coronary arteries during rest. An increased oxygen demand can thus be met only by means of increased circulation. The coronary arteries fill during diastole; the coronary sinus is squeezed during systole. The heart processes glucose, free fatty acids, and lactate. Pulmonary Circulation In the pulmonary circulation, there are regional differences depending on the position of the body: when standing, the basal parts of the lung are better supplied with blood than the apex of the lung. In contrast, however, the apex of the lung is ventilated better: The partial pressure of oxygen in the apex of the lung is 114mmHg, but at the base only 92 mmHg. Euler–Liljestrand Mechanism Low partial pressure of oxygen (a measure of the oxygen content in the air) in an alveolar region results in a contraction of the arterioles in this segment. As a result, circulation is reduced in badly ventilated areas of the lung, but blood flow to the well-ventilated areas increases. If this mechanism causes larger sections of the lung to be taken out of circulation, it can lead to increased resistance in the pulmonary circulation with a greater strain on the right heart. Spinal column deformations such as severe scoliosis can be a cause of this pathology because of reduced chest mobility. Respiratory Regulation Respiratory regulation takes place mainly by chemical control of the carbon dioxide and oxygen content of the blood as well as by measuring the pH level. The carbon dioxide level is therefore the key driving force for respiration: if this value rises in the blood, increased respiration results, to exhale the carbon dioxide. A drop of the pH value to <7.4 has the same effect. The receptors for both of these values are located primarily in the medulla oblongata, where we can also find the respiratory center. The oxygen level is measured in the arch of the aorta and the carotid sinus, and is processed in the medulla oblongata. If the oxygen level drops to about two-thirds of its normal level, increased respiration ensues. The two other measurements, however, respond much earlier with increased respiration. Other respiratory stimuli include:
Anatomy
 Anatomy of the Heart
Anatomy of the Heart
General Facts
Blood Flow in the Bodys
Layered Structure
Impulse-Conducting System
Location
Projection onto the Wall of the Torso
Topographic Relationships
 lung, on both sides
lung, on both sides
 phrenic nerve, on both sides
phrenic nerve, on both sides
 in the pericardial triangle: sternum
in the pericardial triangle: sternum
 ribs 2–6
ribs 2–6
 thymus
thymus
 spinal column
spinal column
 esophagus (bordering on the left atrium)
esophagus (bordering on the left atrium)
 aorta
aorta
 bronchial tubes
bronchial tubes
 pulmonary artery and vein
pulmonary artery and vein
 pulmonary artery and vein
pulmonary artery and vein
 SVC
SVC
 aorta
aorta
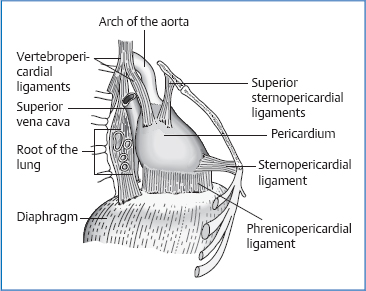
Attachments/Suspensions
 Phrenicopericardial ligaments: in the front right, the pericardium is firmly attached to the diaphragm; elsewhere it can be separated bluntly from the diaphragm.
Phrenicopericardial ligaments: in the front right, the pericardium is firmly attached to the diaphragm; elsewhere it can be separated bluntly from the diaphragm.
 Sternopericardial ligaments: these run from the pericardium to the manubrium and xiphoid process.
Sternopericardial ligaments: these run from the pericardium to the manubrium and xiphoid process.
 vertebropericardial ligaments
vertebropericardial ligaments
 cervicopericardial ligaments
cervicopericardial ligaments
 visceropericardial ligaments
visceropericardial ligaments
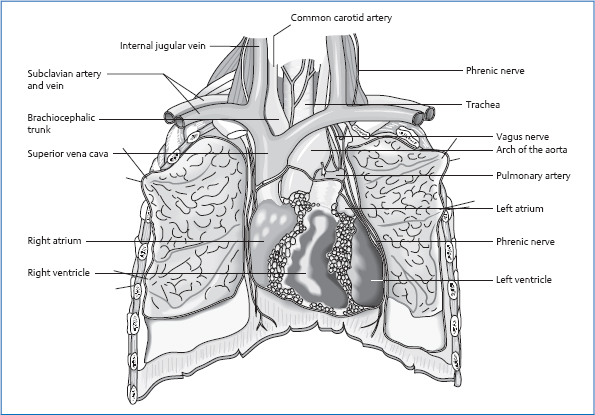
Circulation
Arterial
 left coronary artery:
left coronary artery:
 circumflex branch of the left coronary artery
circumflex branch of the left coronary artery
 both atria
both atria
 right coronary artery:
right coronary artery:
Venous
Lymph Drainage
Innervation
 sympathetic nervous system from T1 to T4
sympathetic nervous system from T1 to T4
 vagus nerve
vagus nerve
 Phrenic nerve supplies the pericardium sensorily together with the sympathetic and parasympathetic nerves.
Phrenic nerve supplies the pericardium sensorily together with the sympathetic and parasympathetic nerves.
Organ Clock
Organ–Tooth Interrelationship
 Anatomy of the Lung
Anatomy of the Lung
General Facts
Location
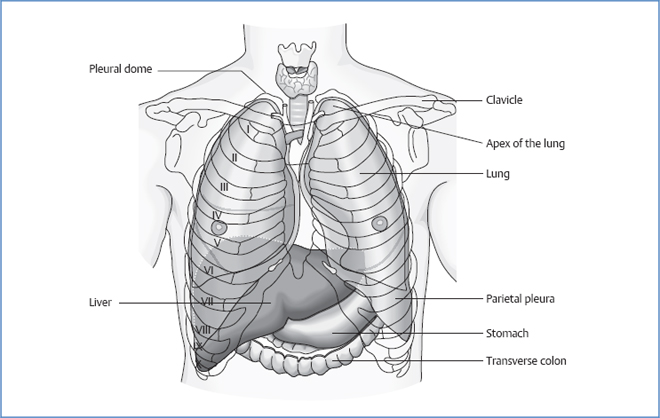
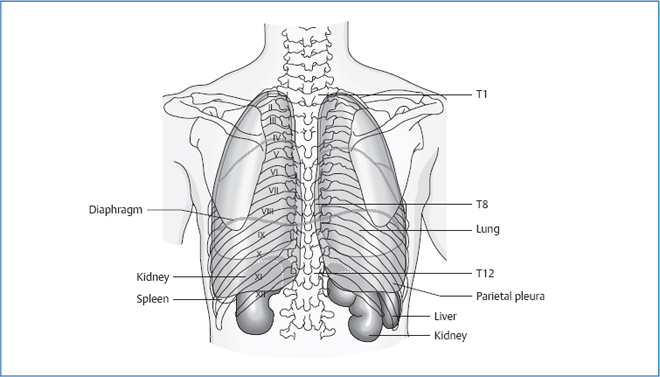
Borders of the Pleura
Borders of the Lung
Location of the Fissures
Topographic Relationships
 ribs 1–12, depending on respiratory position
ribs 1–12, depending on respiratory position
 clavicle
clavicle
 sternum
sternum
 subclavian artery and vein
subclavian artery and vein
 phrenic nerve
phrenic nerve
 pericardiacophrenic artery and vein
pericardiacophrenic artery and vein
 vagus nerve
vagus nerve
 recurrent laryngeal nerve
recurrent laryngeal nerve
 trachea
trachea
 main bronchi
main bronchi
 pulmonary artery and veins
pulmonary artery and veins
 aorta (left)
aorta (left)
 esophagus (right)
esophagus (right)
 heart (more on the left than on the right)
heart (more on the left than on the right)
 diaphragm
diaphragm
 azygos vein
azygos vein
 hemiazygos vein
hemiazygos vein
Attachments/Suspensions
 costopleural ligament
costopleural ligament
 transverse pleural ligament
transverse pleural ligament
 vertebropleural ligament
vertebropleural ligament
Circulation
Arterial
 thoracic aorta for the left lung
thoracic aorta for the left lung
 third and fourth intercostal arteries for the right lung
third and fourth intercostal arteries for the right lung
Venous
Innervation
Organ Clock
Organ–Tooth Interrelationship
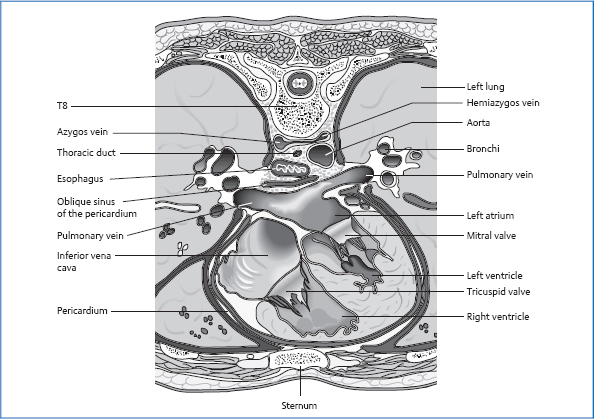
 Anatomy of the Mediastinum
Anatomy of the Mediastinum
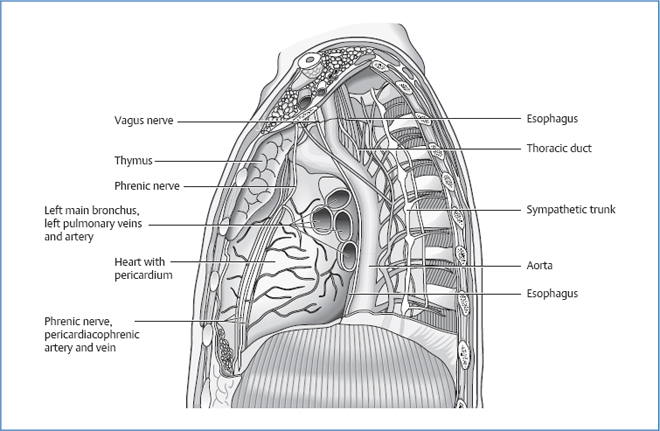
 heart with pericardium
heart with pericardium
 the major arteries and veins of the body:
the major arteries and veins of the body:
 esophagus
esophagus
 trachea
trachea
 main bronchi
main bronchi
 vagus nerve
vagus nerve
 phrenic nerve
phrenic nerve
 sympathetic trunk
sympathetic trunk
 thymus
thymus
 azygos vein
azygos vein
 hemiazygos vein
hemiazygos vein
 thoracic duct
thoracic duct
Movement Physiology
 external intercostals
external intercostals
 serratus posterior superior
serratus posterior superior
 serratus anterior
serratus anterior
 greater pectoral
greater pectoral
 smaller pectoral
smaller pectoral
 sternocleidomastoid
sternocleidomastoid
 erector muscle of the spine
erector muscle of the spine
 abdominal muscles (internal and external oblique, rectus abdominis, transversus abdominis)
abdominal muscles (internal and external oblique, rectus abdominis, transversus abdominis)
 internal intercostal muscle
internal intercostal muscle
 subcostal muscle
subcostal muscle
 transversus thoracis
transversus thoracis
 serratus posterior superior
serratus posterior superior
 latissimus dorsi
latissimus dorsi
Physiology
 Physiology of the Heart
Physiology of the Heart
Systole
Contraction Phase
 ventricle is filled with blood
ventricle is filled with blood
 phase starts when the contraction of the ventricle starts
phase starts when the contraction of the ventricle starts
Ejection Phase
Diastole
Relaxation Phase
Fill Phase
Heart Sounds
 at the start of systole
at the start of systole
 The isovolumetric contraction of the ventricles causes the ventricles and AV valves to vibrate. This creates a sound.
The isovolumetric contraction of the ventricles causes the ventricles and AV valves to vibrate. This creates a sound.
 at the start of diastole
at the start of diastole
 The closure of the semilunar valves creates the second heart sound.
The closure of the semilunar valves creates the second heart sound.
Dynamics of the Heart—Adaptation to Changing Strains
Acute Volume Overload
Acute Pressure Overload
Energetics of the Heart
 Physiology of the Lung
Physiology of the Lung
 muscle activity
muscle activity
 warmth or cold
warmth or cold
 changes in body temperature
changes in body temperature
 pain
pain
 epinephrine and progesterone
epinephrine and progesterone
 drop in blood pressure
drop in blood pressure
Pathologies
Symptoms that Require Medical Clarification
Related posts:


Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


