CHAPTER 9 Role of oxytocin and oxytocin-related effects in manual therapies
The relaxation and growth or the calm and connection response
All types of manual therapy involve touching the patient. In this chapter we suggest that all therapies involving gentle skin contact will lead to activation of a basic psychophysiological reaction in response to activation of cutaneous sensory nerves. This basic reaction pattern is probably from an evolutionary perspective very old, and forms an antithesis to the fight and flight response and other defensive reactions (Cannon 1929, Selye 1976). The effects include stimulation of social behavior, decreased anxiety, increased pain threshold, wellbeing, decreased activity in the hypothalamopituitary–adrenal (HPA) axis, lowered blood pressure, vasodilatation of cutaneous vessels, and increased function in the endocrine system of the gastrointestinal tract. In addition, anabolic, storage metabolism, healing of wounds and restorative processes are promoted. Learning is also stimulated.
Corticotropin-releasing factor (CRF) and vasopressin play an important integrative function in defense and stress reactions. We propose that oxytocin produced in the nucleus paraventricularis (PVN), mainly via oxytocinergic nerves projecting to many important regulatory areas in the central nervous system (CNS), plays a similar integrative function in a reaction pattern that runs opposite to and forms an antithesis to the defense reactions. The physiological aspects of this reaction pattern have been named the relaxation and growth response and, when behavioral expressions are included, the calm and connection response (Uvnäs-Moberg 1997, 2003).
Noxious versus non-noxious stimulation of sensory nerves and the experience of fear versus trust
Release of CRF and vasopressin can be induced by physical damage to the skin or other noxious stimuli by activation of cutaneous sensory nerves. In contrast, oxytocin is released in response to non-noxious sensory stimulation of the skin, e.g., by touch, stroking, and warmth. In addition, mental experiences of danger or threat can, via processing in the hippocampus and amygdala axis, induce stress responses and defense. By analogy, we suggest that the oxytocin release and oxytocin-related effect patterns can be induced by mental experiences of love, trust, and safety (Uvnäs-Moberg et al. 2005).
The role of touch and trust in manual therapies
Sensory nerves mediating effects of non-noxious sensory stimulation
Recently it has been demonstrated that also a subgroup of the thinner and more slowly conducting C fibers (CT = C tactile) mediates pleasant feelings and can be activated by touch, not only in animals but also in humans. From an evolutionary perspective, CT fibers are older than the thicker and more rapidly conducting A and B fibers, and are mainly connected to older parts of the brain, where they influence emotions. The CT fibers are activated especially in response to slow and light stroking (around 40 cm/min) and have been shown to activate areas in the insular cortex, which are related to the sensations of wellbeing (Craig 2003, Olausson et al. 2002, Vallbo et al. 1999).
Effects of non-noxious sensory stimulation in animals
Noxious stimulation is well known to induce both defense reactions and physiological reactions, which are compatible with activity and stress, via an increased activity in the HPA axis, relevant aspects of the sympathoneural system and of the sympathoadrenal (SA) system. Non-noxious stimulation, however, also induces effects of physiological nature. In anesthetized rats brushing or stroking reduces activity in the SA system and in the HPA axis, as evidenced by lowered levels of adrenaline and of corticosterone in the circulation (Araki et al. 1984, Kurosawa et al. 1982, Tsuchiya et al. 1991). Blood pressure is also reduced (Kurosawa et al. 1995). In addition, efferent vagal nerve activity is increased by low-intensity electrical stimulation and by stroking, as evidenced by an influence on the levels of vagally controlled gastrointestinal hormones (Uvnäs-Moberg et al. 1992).
Behavioral effects are also induced by stroking: for example it induces a sedative effect by reducing spontaneous motor activity. This effect is dependent on the amount of stroking given, as no effect was observed after 2 minutes of stroking but was seen after 5 minutes. Weight gain is observed in response to repeated treatments with stroking, and the increase in nociceptive thresholds is further enhanced (Ågren et al. 1995, Holst et al. 2002, 2005, Kurosawa et al. 1995, Lund et al. 1999, 2002, Uvnäs-Moberg et al. 1992, 1993, 1996a).
Effects of non-noxious sensory stimulation can also be observed as a consequence of maternal interaction with pups. Stress-related effects in rat pups, such as elevated levels of corticosterone and decreased levels of growth hormone as a consequence of separation from their mothers, are prevented if the pups are exposed to extra stroking (Pauk et al. 1986, van Oers et al. 1998). The mothers are also kept calm when they are in close contact with their offspring (Lonstein 2005).
Stroking of rat pups may even induce lifelong effects. When pups were exposed to daily stroking on the ventral side of the abdomen for 1 week postnatally, blood pressure was significantly reduced in adulthood (Holst et al. 2002). Further, rat pups exposed to mothers that are highly interacting during the first week of life become less anxious, more social, and more tolerant to stress than pups of mothers who interact less (Champagne & Meaney 2008).
Oxytocin
Oxytocin – a neurotransmitter and a hormone
Oxytocin is a nonapeptide produced in the PVN and the supraoptic nucleus (SON) with- in the hypothalamus. Magnocellular neurons within the SON and PVN project to the neurohypophysis, whence oxytocin is released into the circulation. In addition, a widespread network of parvocellular, oxytocinergic nerve fibers project from the PVN to many areas within the brain. The median eminence, the amygdala, the hippocampus, the olfactory bulb, the striatum, the nucleus accumbens (NA), the raphe nuclei, the locus ceruleus, the vagal motor and sensory nuclei, and the spinal cord are all reached by oxytocinergic fibers (Buijs et al. 1985).
Oxytocin may induce effects in many areas of the brain, as oxytocin receptors are widely distributed in the brain (Freund-Mercier et al. 1987). Recently it has been demonstrated that large quantities of oxytocin are also released from cell bodies and dendrites of oxytocin-producing cells in the SON and PVN. It has therefore been suggested that oxytocin may also reach neurons that are not adjacent to the release sites by diffusion or volume transmission (Ludwig & Leng 2006).
It is well known that the mammary glands and the uterus are richly provided with oxytocin receptors. Oxytocin receptors are, however, also found in many peripheral organs, e.g. in the kidneys, pancreas, stomach, thymus, adipocytes, heart, ventricle and blood vessels (Bonne & Cohen 1975, Elands et al. 1990, McCann et al. 2002, Stock et al. 1990, Stoeckel & Freund-Mercier 1989, Yazawa et al. 1996).
The PVN is also reached by afferent pathways from the NTS, the locus ceruleus, other parts of the hypothalamus, and even from the dorsal horn of the spinal cord, suggesting that the release of oxytocin is controlled from many different areas in the CNS (Swanson & Sawchenko 1980, 1983, Sawchenko & Swanson 1982).
The distribution of afferent and efferent projection patterns and the distribution of oxytocin receptors are similar in females and males (Sawchenko & Swanson 1982, Sofroniew 1983, Swanson & Sawchenko 1980, 1983), but the effects of oxytocin may differ between males and females owing to the effects of sex hormones. Estrogens may increase the release of oxytocin and also the responsiveness and/or number of oxytocin receptors (Schumacher et al. 1993).
Effects of oxytocin in animals
Oxytocin administered intracerebroventricularly, or in high doses systemically to induce effects in the CNS, has been demonstrated to reduce blood pressure and activity in the HPA axis. The effect of oxytocin on the HPA axis seems to be exerted at many levels (see below and Fig. 9.1). It also influences the release of gastrointestinal hormones, increases nociceptive thresholds, and reduces inflammation and levels of anxiety, as well as inducing calm and stimulating learning.
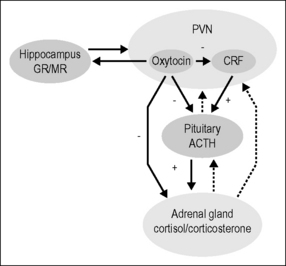
When oxytocin is administered repeatedly long-term effects may be induced. Five daily injections of oxytocin reduce blood pressure and activity in the HPA axis and increase nociceptive thresholds for weeks after the last treatment. In response to repeated treatments oxytocin also influences the levels of gastrointestinal hormones, increases weight gain, and shortens wound healing time. Further, it improves learning by conditioning (Björkstrand et al. 1996, Petersson et al. 1996a,b, 1998, 1999a,b, 2001, Szeto et al. 2008, Uvnäs-Moberg et al. 1992, 1996b, 2000).
Oxytocin knockout mice have an increased release of corticosterone in response to stress and show more anxiety-like behavior, further supporting the role of oxytocin as an anxiolytic and anti-stress agent (Amico et al. 2008). In addition, oxytocin stimulates various kinds of interactive social behavior and bonding between mother and young, and between males and females (Carter 1998).
Oxytocin mediates effects not only via the classic uterine oxytocin receptor, but proba-bly also via oxytocin receptors with other characteristics, as not all oxytocin effects exerted in the brain are blocked by antagonists directed towards the classic oxytocin receptor. Moreover, administration of oxytocin to rats increases α2-adrenoreceptor function in several brain regions, such as the amygdala, NTS, and locus ceruleus, as measured by autoradiography and electrophysiology. It is likely that this effect underlies some of the oxytocin-induced anti-stress effects (Petersson et al. 2005). Oxytocin also influences cholinergic, serotonergic (5HT), and opioidergic transmission. Some of the long-term effects may be induced by altering the function of these classic signaling systems. In addition, dopamine, opioids, acetylcholine, and 5HT influence the release of oxytocin, and antidepressant drugs, such as serotonin reuptake inhibitors (SSRI), have been shown to release oxytocin (Uvnäs-Moberg et al. 1999).
Taken together, these data show, first, that oxytocin induces a response pattern cha- racterized by anti-stress effects and stimulation of growth and restorative processes (the growth and relaxation response) and also behavioral actions involving calm and increased social behavior (the calm and connection response) (Uvnäs-Moberg 1997, 2003); and second, that the function of the oxytocinergic system is strongly connected with the activity of other, more classic, signaling systems (Bagdy & Kalogeras 1993, Clarke et al. 1978, Crowley et al. 1991, Uvnäs-Moberg et al. 1999, Wright & Clarke 1984).
Effects of oxytocin in humans
Since the beginning of the 19th century oxytocin has been known to stimulate uterine contractions during labor and to induce milk letdown in breastfeeding women, being administered as an infusion to induce or augment labor and as a nasal spray to promote milk letdown. Recently the oxytocin nasal spray has also been administered to men, and several effects have been documented. The oxytocin has an anxiolytic effect and dampens activity in the amygdala. It also reduces activity in the HPA axis, as well as increasing social skills, e.g. the ability to read and evaluate the emotional valence of faces, in both healthy and autistic men. In other studies the administration of oxytocin spray has been demonstrated to increase trust and generosity in men and to reduce abdominal pain in women (Heinrichs et al. 2003, Kosfeld et al. 2005, Ohlsson et al. 2005, Rimmele et al. 2009). Oxytocin administered as an intravenous infusion also reduces anxiety and increases social skills, effects that last for weeks after administration (Hollander et al. 2007, Jonas et al. 2008a).
Oxytocin release in response to non-noxious sensory stimulation in animal experiments
It is well known that oxytocin is released during parturition and in response to suckling in lactating animals, and in response to sexual interaction. Oxytocin can, however, also be released in response to other types of non-noxious and pleasant sensory stimulation. When anesthetized rats were exposed to gentle stroking on their backs or afferent electrical stimulation of the sciatic or the vagal nerves, oxytocin levels in plasma increased more than twofold (Stock & Uvnäs-Moberg 1988), and when they were exposed to electroacupuncture, thermal stimulation, or vibration, oxytocin levels increased in both plasma and cerebrospinal fluid (Uvnäs-Moberg et al. 1993).
As mentioned above, 5 minutes of stroking on the abdomen of conscious rats (40 strokes per minute) induces a multitude of behavioral and physiological effects (Box 9.1; note that weight gain is only observed in response to repeated treatments). Oxytocin is also released by this type of stimulation, and the effects of stroking may therefore involve an oxytocinergic mechanism. The elevation of pain threshold and the calming and anxiolytic-like effects caused by stroking on the abdomen are probably exerted in the periaqueductal gray (PAG), amygdala, and LC, by oxytocin released from oxytocinergic fibers originating in the PVN. The connection between oxytocin and the effects induced by stroking are supported by findings that some of the effects, e.g. the elevation of pain threshold, caused by the stroking treatment are blocked if the animals are given an oxytocin antagonist beforehand. In further support of a role for oxytocin in the increase in pain threshold caused by stroking is the fact that increased oxytocin levels in response to stroking have been demonstrated in the PAG, an area in the brain that is of central importance for nociception (Lund et al. 2002).
Box 9.1 Effects of stroking
Sedative and anxiolytic-like effects
Increased nociceptive thresholds
Decreased heart rate and blood pressure
Changed levels of cholecystokinin, gastrin, and insulin
It is important to state that the effects induced by oxytocin on physiological and behavioral function in response to sensory stimulation are exerted in the brain. Therefore, circulating levels of oxytocin are not always a relevant marker for the release of oxytocin in the brain. Oxytocin may be released into the brain in many situations without a concomitant release into the circulation. A parallel secretion of oxytocin into both the brain and into the circulation has, however, been shown during suckling, feeding, parturition, and vaginocervical stimulation (Kendrick et al. 1986, 1988). In rats, electroacupuncture, vibration, and thermal stimuli significantly increase oxytocin levels in the cerebrospinal fluid (Uvnäs-Moberg et al. 1993).
Related posts:
 Manual medicine and the autonomic nervous system: assessing autonomic function in humans
Manual medicine and the autonomic nervous system: assessing autonomic function in humans
 A physical therapist’s perspective on manual therapy: clinical effectiveness and selected mechanism
A physical therapist’s perspective on manual therapy: clinical effectiveness and selected mechanism
 Survey of mechanotransduction disorders
Survey of mechanotransduction disorders
 Viscerosensory pathways in the brain
Viscerosensory pathways in the brain
 Research related to clinical applications of manual therapy for musculoskeletal and systemic disorders from the osteopathic experience
Research related to clinical applications of manual therapy for musculoskeletal and systemic disorders from the osteopathic experience
 Spinothalamic system and viscerosomatic motor reflexes: functional organization of cardiac and somatic input
Spinothalamic system and viscerosomatic motor reflexes: functional organization of cardiac and somatic input
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


