CHAPTER 1 Phylogeny
EVOLUTION OF THE VERTEBRATE ELBOW
The distal humerus of pelycosaurs, the late Paleozoic (255 to 235 mya) reptiles that probably gave rise to more advanced mammal-like reptiles, possessed a bulbous capitellum laterally and medially. The articulation with the ulna was formed by two distinct surfaces: a slightly concave ventral surface and a more flat dorsal surface (Fig. 1-1).11 The proximal articular surface of the ulna was similarly divided into two surfaces separated by a low ridge. Reconstruction of the forelimb of these reptiles suggests that they walked with limbs splayed out to the side. The humerus was held more or less horizontal, the elbow flexed to 90 degrees, and the forearm was sagittally oriented. Forward motion was brought about by rotation of the humerus around its long axis, which propelled the body forward relative to the fixed forefoot. Elbow flexion and extension probably were useful only in side-to-side motions. The ulnohumeral joint, with its dual articular surfaces, was well suited to resist the valgus/varus stress produced by humeral rotation, and the proximal end of the radius was flat and triangular, precluding pronosupination. It appears, therefore, that stability rather than mobility was the major functional characteristic of the elbow of these late Paleozoic reptiles.
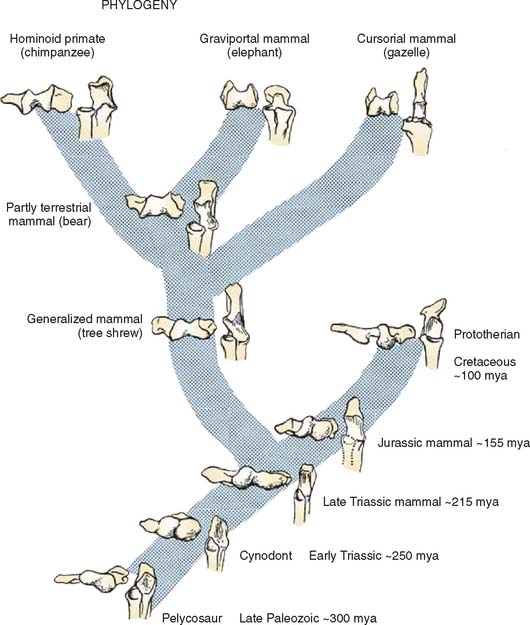
Cynodonts, the more immediate ancestors of mammals from the Permo-Triassic period (235 to 160 mya), had their limbs underneath their bodies rather than at the sides. The distal humeral articular surface consisted of radial and ulnar condyles separated by a shallow groove (see Fig. 1-1). The proximal ulnar articular surface was an elongate spoon shape for articulation with the humeroulnar condyle. The lateral flange on the ulna for articulation with the radius was separated from this surface by a low ridge. This ridge articulated with the groove between the radial and ulnar condyles displaying some features in common with the “tongue and groove” (trochleariform) type of humeroulnar articulation characteristic of many modern mammals.
Early mammals from the Triassic (210 to 160 mya) and Jurassic (160 to 130 mya) periods also had radial and ulnar condyles. However, the radial condyle was more protuberant than the ulnar, and the ulnar condyle was more linear and obliquely oriented (see Fig. 1-1). The two condyles were separated by an intercondylar groove. The ulnar notch had articular surfaces for both the ulnar and the radial condyles, each matching the configuration of the corresponding humeral surface. The oblique orientation of the humeroulnar joint resembled a spiral configuration, which helped to keep forearm movement in a sagittal plane as the humerus underwent a compound motion involving adduction, elevation, and rotation during propulsion.
The trochleariform distal humeral articular surface in modern mammals largely came about by widening the intercondylar groove and the development of a ridge within it (see Fig. 1-1, bear). The articular surface on the proximal ulna is oblique in orientation, and the distal half retains an articulation with the ulnar condyle. This spiral trochlear configuration allows the forearm to move in a sagittal plane while maintaining the stability of ulnohumeral contact through the cam effect of the ulnar condyle during humeral rotation.
COMPARATIVE PRIMATE ANATOMY OF THE ELBOW REGION
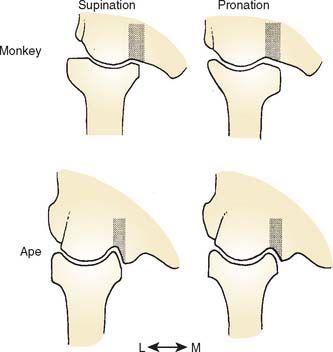
Much of what follows is taken from the detailed studies of Rose.20,21 The humeral trochlea may be cylindrical, conical, or trochleariform in nonhuman primates.21 The trochlea is conical in some prosimians, but a cylindrical trochlea seems to be the most common shape and is observed in most prosimians and New World monkeys. The trochlea is also cylindrical in most Old World monkeys but with a pronounced medial flange or keel that is best developed anterodistally (Fig. 1-2). Only in apes and humans is the trochlea truly trochleariform, possessing medial and lateral ridges all around the trochlear margins, which contribute to the stability of the ulnohumeral joint, substituting for the radiohumeral joint, which is freed for pronosupination throughout the flexion range.11,20 In most species, the articular surface of the trochlea expands posteriorly to the area behind the capitellum. In larger monkeys, the lateral edge of the posterior trochlear surface projects to form a keel that extends up the lateral wall of the olecranon fossa (see Fig. 1-2). In hominoids, this keel is a continuation of the lateral trochlear ridge and helps form a sharp lateral margin of the olecranon fossa, providing resistance to varus and internal rotation in extension.20.21
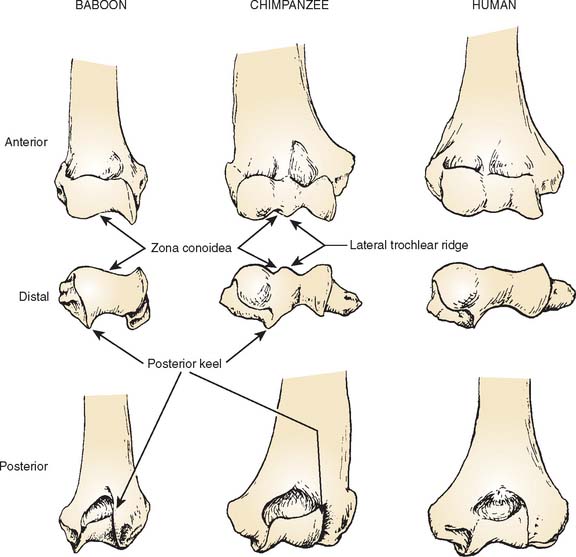
The trochlear notch of the ulna generally mirrors the shape of the humeral trochlea. In humans and apes, the notch has medial and lateral surfaces separated by a ridge that articulates with the trochlear groove (Fig. 1-3).20,21
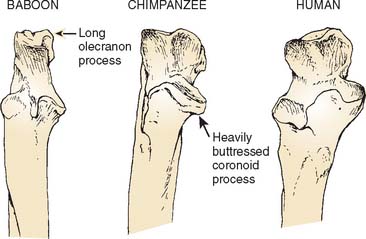
The differences seen in the configuration of the humeroulnar joint across primate species reflect contrasting requirements for stabilization with different forms of limb use. In most monkeys, the humeroulnar joint is in its most stable configuration in a partially flexed position owing to the development of the medial trochlear keel anterodistally and the lateral keel posteriorly.20
The great apes (chimpanzees, gorillas, and orangutans) and the lesser apes (gibbons) move about in a much less stereotypical fashion than do monkeys. To accommodate this more varied form of limb use, the hominoid humeroulnar joint, with its deeply socketed articular surfaces and well-developed medial and lateral trochlear ridges all around the joint margins, is designed to provide maximum stability throughout the flexion-extension range.20–22 The use of overhead suspensory postures and locomotion in apes has led to the evolution of the capacity for complete elbow extension. Apes even keep their elbows extended during quadrupedal locomotion. The ideal joint configuration for resistance of transarticular stress with fully extended elbows during quadrupedal postures would be to have a trochlear notch that was proximally directed. It could then act as a cradle to support the humerus during locomotion. However, a proximal orientation of the trochlear notch would severely limit elbow flexion by impingement of the coronoid process within its fossa. The anteroproximal orientation of the trochlear notch in apes thus represents a compromise that safely supports the humerus on the ulna in extended elbow positions during locomotion without unduly sacrificing elbow flexion.1
On the lateral side of the elbow, the articular surface on the capitellum extends farther posteriorly in apes and humans than in monkeys, allowing the radius to move with the ulna into full extension of the elbow. In addition, the capitellum of apes and humans is uniformly rounded, reflecting versatility rather than stereotypy in forelimb usage (Fig. 1-4).
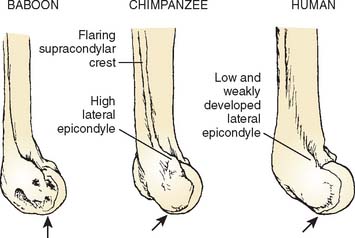
The gutter-like region between the trochlea and capitellum—the zona conoidea—is a relatively flat plane that terminates distally in most monkeys. In the hominoids, it continues posteriorly (see Fig. 1-1).20,21 The zona conoidea articulates with the bevel of the radial head, and differences in its configuration reflect differences in the shape of the radial head.
The radial head of hominoid primates is nearly circular, and the peripheral rim is symmetrical and beveled all around the circumference of the radial head for articulation with the zona conoidea (Fig. 1-5). This configuration provides good contact to resist dislocation of the radial head from the humerus under the varied loading regimes experienced by the hominoid elbow and can stabilize the radial head in all positions of pronosupination.20,21

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


