Chapter 2 Functional Anatomy of the Cervical Spine
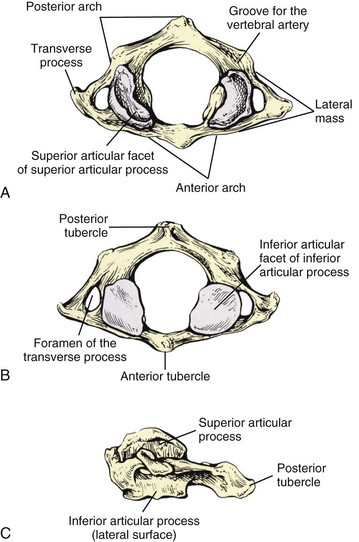
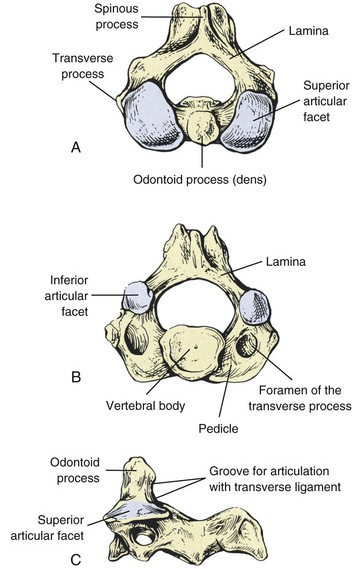
In the cervical region, normal movement, as in all regions of the spine, is dependent on spinal motion segments1 (Table 2-1). In the lower cervical spine, the motion segments are typical of the spine in general, whereas the three upper cervical motion segments are atypical.2 The typical motion segment is made up of two vertebral bodies, the intervening intervertebral disc, two posterior spinal joints, neurological elements confined within the two lateral recesses and intervertebral foramina, plus all the connective and muscular tissues supporting and limiting intersegmental movement (Figure 2-1, Table 2-2). The upper two cervical motion segments provide for a wide range of movement (Figures 2-2, 2-3, 2-4) even though no intervertebral discs separate the anterior portions of these segments. The occipito-atlantal articulation (C0-C1) has the two paired condyles of the occiput, which fit into the concave articular surfaces of the atlas (Figure 2-3A). The body of the atlas in the atlantoaxial (C1-C2) segment is replaced anteriorly with the peg-like odontoid process of the axis. (See Figures 2-3B, 2-3C, 2-4.) This process is bounded anteriorly by the anterior arch of the atlas and posteriorly by the transverse cruciate ligament. Despite the lack of intervertebral discs that allow six degrees of freedom of movement in the typical motion segment, the two atypical upper spinal motion segments contribute to a considerable amount of movement in the upper cervical region. (See Table 2-1.) The C0-C1 articulation allows for cervical flexion and extension, while the C1-C2 motion segment accounts for 50% of the total cervical rotation. However, the increased demand for motion in the cervical spine affects its strength properties. In comparison to the lumbar spine, the cervical spine has approximately 20% of its bending strength and 45% of its compressive strength, making it more vulnerable to bending injuries.3 The joints of Luschka (uncovertebral joints) and the transverse foramen through which the vertebral arteries pass are peculiar to the cervical spine.
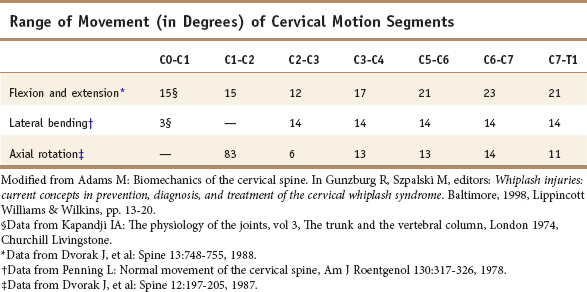
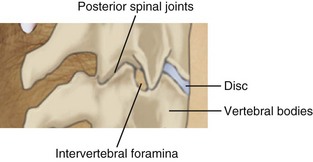
TABLE 2-2 Primary Function of the Components of the Lower Cervical Spinal Motion Segments
| Vertebrae | Support spinal column and protect spinal cord |
| Intervertebral discs | Unite vertebrae and provide shock absorption |
| Zygapophyseal joints | Guide and restrict motion |
| Anterior and posterior longitudinal ligaments | Connect vertebrae and support spinal column |
| Ligamentum nuchae | Connects laminae and protects spinal cord |
| Z joint capsules | Reinforces the zygapophyseal joints |
| Intertransverse, supraspinous, and interspinous ligaments | Add stability to the spinal column |
| Transversospinalis muscles | Segmental position sensors |
| Short segmental muscles | Segmental position sensors |
| Uncovertebral joints | Restricts lateral flexion |
Embryological Development of the Spinal Motion Segments
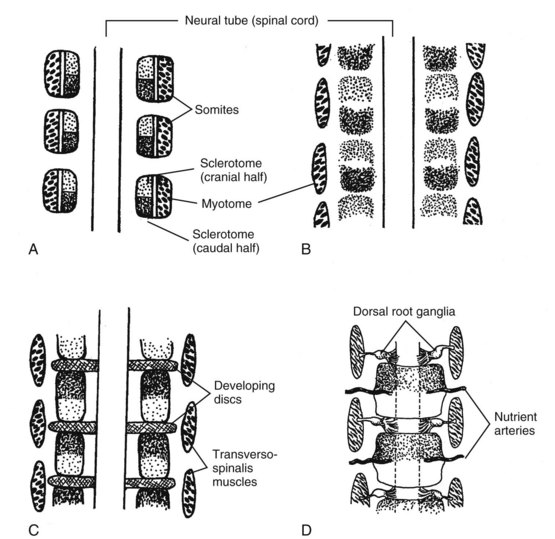
The vertebrae of the spinal column are derived from mesenchyme, which subsequently undergoes chondrification and ossification.4 Before reaching its final condition, the spinal column passes through four major stages: the nonsegmental axis, the segmented mesoderm, chondrification, and ossification (Box 2-1). The major embryological stages leading to the development of the spinal column and associated structures involve a recombination of the somites that allows for segmental spinal motion (Figure 2-5A-D).
Somatogenesis
Somites begin formation as discrete clusters of mesenchymal cells that undergo segmentation in a cranial to caudal progression beginning around the third week of intrauterine life; this stage is termed “compaction.” A single somite can be described as having six faces, like a cube, with each facet having a slightly different fate. Further, the position along the embryo can alter the developmental fate of the somite. During this stage, mesenchymal cells multiply rapidly, enclosing the notochord ventromedially as segmented sclerotomes. (See Figure 2-4A.) These will give rise to the bones, joints, and ligaments of the spinal column. Sclerotomal cells later migrate dorsally around the spinal cord, forming the neural arches. The mesenchymatous models in the third stage produce a cartilaginous vertebral column. Single mesenchyme cells will differentiate into chondroblasts when they are maintained in a rounded configuration. In the fourth stage of spinal development, ossification of the cartilaginous vertebral column occurs.4
The segmental nature of the mature vertebral column reflects its origin from the mesenchymal somites of the early embryo. During the fourth week, the ventromedial wall of each somite separates into a mass of diffuse cells, forming the sclerotomes from which the vertebrae originate.4 The cells of the dorsolateral part of the somite make up the dermomyotome. The dermatome or skin plate lays the foundation for the dermis, and the myotome or muscle plate is the forerunner of striated muscle. This differentiation becomes clinically significant to sclerotomal and dermatomal pain patterns and their origin, corresponding to the development of the spinal nerves directly opposite their respective somites.
The sclerotomal cells from each somite pair are densely packed in the caudal region and loosely packed in the cranial region. (See Figure 2-4A.) As development proceeds, a sclerotomic fissure separates these areas (see Figure 2-4B), and the component halves of adjacent sclerotomes reunite in new combinations. (See Figure 2-4B.) The denser caudal section of each original sclerotome joins the looser cranial half of the adjacent caudal sclerotome. (See Figure 2-4C.) These recombinations then become the primordia of the vertebrae. Their fusion around the notochord produces the vertebral centrum (body). The cranial portion of the condensed caudal mass, which is farthest from the nutrition provided by the artery, increases in density, forming the precursor of the intervertebral disc. The poor vascularity of the early intervertebral disc is lost in adult life, and the nutritional demands are met by diffusion from lymph.
The mesenchymal cells located between the cephalic and caudal portions of the original sclerotome segments fill the space between the two precartilaginous vertebral bodies as precursors of the intervertebral discs.4 Whereas the sclerotomal mesenchyme forming the body of the vertebrae replaces the notochordal tissue that it surrounds, the notochord expands as localized aggregates of cells and matrix, thus forming the nucleus pulposus of the intervertebral disc where it persists until the second decade. The notochord then undergoes mucoid degeneration that forms the noncellular matrix of the nucleus pulposus. The nuclear material after 10 embryonic weeks is surrounded by the circular fibers of the annulus fibrosis, which is formed into a laminated fibrous zone from fibroblastic cells. These two structures form the intervertebral disc, giving it a double origin: the central nucleus pulposus from the notochord, and the annulus from the fibroblastic extension of the vertebral bodies.
The skeletal muscles develop from the dorsolateral or myotomal portion of the somite. When the vertebral bodies recombine, the adjacent myotomes that do not split now span the intervertebral fissure. Thus, the myotome-derived muscles are in a position to move, monitor, or stabilize adjacent vertebrae relative to each other. (See Figure 2-5.) This mechanical relationship establishes the elements of the spinal motion segments defined as two adjacent vertebrae and the connecting tissue binding them to each other.5 With recombination of the sclerotomal portions, the intersegmental arteries originally located between the sclerotomes eventually pass into the middle of the vertebral bodies. (See Figure 2-5.)
Dorsal and lateral to the developing centra (body of the vertebra), the less dense portions of the sclerotomes form the precartilaginous neural arches and transverse processes, while denser portions give rise to the intervertebral ligaments.4 Segmentation seems to be induced by the pull of the notochord, which acts as an organizer in effecting the centrum. The neural crest cells that develop into the dorsal root ganglia appear to act as organizers for vertebral arch segmentation.
Components of the Lower Spinal Motion Segments
Vertebrae
The lower cervical vertebrae have three parts that are associated with movement: the anterior (ventral) body, the medial (neural) arch, and the lateral transverse and posterior (dorsal) spinous processes (Figures 2-6A, 2-6B). All three parts have a predominant function in addition to sharing common functions. The anterior portion serves mainly as a load distributor. The medial portion serves to guide and restrict motion that is dependent on the geometry, size, and spatial orientation of the articular facets (articular processes). The combined anterior and medial portions provide necessary protection to the spinal cord, meninges, and their vessels. The lateral transverse and posterior spinous processes function as lever-like extensions for the attachment of muscles and the stabilizing ligaments. The foramen of the transverse processes provides openings through which the vertebral arteries pass upward through the transverse processes.
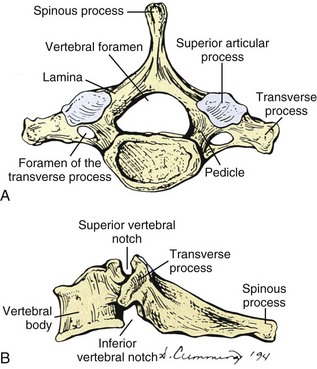
Figure 2-6 A, Superior and B, Lateral views of the seventh cervical vertebra, the vertebral prominens.
All three parts of the vertebrae interact for normal vertebral movement to occur.
The complete column of bodies and discs forms a strong but flexible central axis. The intervertebral discs provide for stability and movement, as connectors between vertebrae, and as separators that function as deformable spacers that allow for movement. The intervertebral foramen between adjoining neural arches near their junctions with the vertebral bodies allows passage of mixed spinal nerves, smaller recurrent nerves, and blood and lymphatic vessels.6
The vertebral bodies are convex anteriorly and concave posteriorly where they complete the vertebral foramen. The vertebral arch has on each side a vertically narrow ventral part, the pedicle, and dorsally a broader lamina. Adjacent vertebral notches contribute to the intervertebral foramen when the three-joint complex (disc and two zygapophyseal joints) articulates.6 The side-to-side diameter of the cervical vertebrae increases from C2 to C7 to allow the lower vertebrae to support the greater weight that they are required to carry. The anterior surfaces of the vertebral bodies can develop osteophytes (bony spurs). Asymptomatic osteophytes that occur in 20% to 30% of the population can become clinically significant with injury to the cervical spine. Swelling that causes pressure on the more anteriorly located esophagus or trachea may lead to difficulty swallowing (dysphagia) and difficulty with speech (dysphonia). The superior and inferior surfaces of the vertebral bodies are described as being sellar or saddle shaped. The superior surface is concave from left to right as a result of the raised lateral lip. The superior surface is convex from front to back because of the beveling of the anterior surface.
Intervertebral Discs
The intervertebral discs of the cervical spine make up more than 20% of the superior-inferior length of the cervical spine. These structures allow the large amount of movement that occurs in the cervical region. Intervertebral discs are situated between adjacent surfaces of vertebral bodies between C2 (axis) and C3 through C7 and T1. The discs provide almost 40% of the height of the cervical spine from C2 to C7. The C2-C3 interbody joint is the most superior disc capable of affecting a spinal nerve (C3 spinal nerve).7
Each intervertebral disc is a fibrocartilaginous coupling that forms an articulation between the bodies of the vertebrae. It serves both to unite the adjacent vertebral bodies and to hold them apart by means of the hydrostatic pressure of the centrally located nucleus pulposus. The nucleus exhibits considerable elastic rebound that allows the disc to assume its original physical state upon release of pressure. Recently, elastin content in the disc has been studied. Approximately 2% of the disc is made up of elastin, with the outer annulus fibrosus, inner annulus fibrosus, and nucleus pulposus having similar levels of elastin. Although the exact function of elastin in the healthy disc is unknown, degenerated discs exhibited a significant increase in elastin content, particularly in the inner annulus fibrosus. The increase in elastin content is thought to be in response to the deformation of the lamellar structure during radially applied loads, thereby resisting delamination.8
Mercer and Bogduk9 demonstrated that the annulus fibrosus of the cervical disc is different than that of the lumbar spine. Instead of a ring of concentric lamella, the annulus fibrosus is arranged in a crescent-shaped mass of collagen that is thick anteriorly and tapers toward the uncinate processes laterally (Figure 2-7). The annulus fibrosus is deficient posterolaterally and posteriorly; only a thin vertical layer of annular fibers is present. (See Figure 2-7.) Mercer and Bogduk9 concluded that the annulus fibrosus was more like an interosseous ligament than a ring of annular fibers around the nucleus pulposus. The difference in morphology of the cervical disc may help to explain some of the difference in biomechanics of the cervical spine related to the lumbar or thoracic spine.
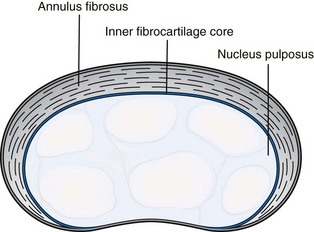
Figure 2-7 Superior view of a cervical intervertebral disc illustrating the crescent-shaped annulus fibrosus.
In the adult nucleus pulposus there is a progressive increase in the collagen content with a corresponding decrease in the cell population. The sparser cell population associated with reduced vascularity continues to produce proteoglycans. The peripheral annulus contains a few small blood vessels. Normal adult discs have a high level of hydration and can absorb water readily, particularly in the nucleus. Disc turgor is created by the high water content that is maintained by the enveloping cartilaginous envelope. The nucleus pulposus is separated from the central parts of the vertebral bodies by only thin cartilaginous plates. At the centers of these, where the notochord originally penetrated the cartilage plates, there are weak areas where sudden axial loading of the spine may cause herniation of the young turgid nucleus into the vertebral spongiosa. These depressions in the vertebral end plates are called Schmorl’s nodes and are found commonly in children and over a third of all adults. They apparently produce little if any adverse effect on the functioning of the intervertebral disc.10
The intervertebral disc has been found to have both vasomotor and sensory innervation.11 The vasomotor fibers are associated with the small vessels located along the superficial aspect of the annulus fibrosis. The sensory fibers are thought to be both nociceptive (pain sensitive) and proprioceptive. Mendel et al12 found sensory fibers throughout the annulus fibrosis in cervical intervertebral discs. They were most numerous in the middle third of the disc (superior to inferior) and were consistent with those that transmit pain. In addition, pacinian corpuscles and Golgi tendon organs were found in the posterolateral aspect of the disc. The posterior aspect of the disc receives its innervation from the recurrent meningeal (sinuvertebral) nerve. The posterolateral aspect of the annulus receives both direct branches from the anterior primary division and also branches from the gray communicating rami of the sympathetic chain. The lateral and anterior aspects of the disc primarily receive their innervation from branches of the gray communicating rami and also from branches from the sympathetic chain. It does not appear that nerve fibers are present in the nucleus pulposus.12
Disc degeneration occurs with aging and is manifested by a complex process of changes. The disappearance of notochordal cells in the first decade of life from the nucleus pulposus is followed by loss of its gel-like structure from a decrease in cells, water, and proteoglycans from the extracellular matrix. The annulus fibrosus also loses its shape, which provides mechanical strength.13
Posterior Spinal (Zygapophyseal) Articulations
The posterior spinal articulations, commonly referred to as the zygapophyseal joints, are true diarthrodial joints with characteristics similar to those of peripheral diarthrodial joints (Figure 2-8). They have articular cartilage, a loose capsule lined with synovial membrane, reinforcing ligaments, and related muscles. They appear on both sides of the posterior aspect of the lower cervical motion segments, guiding and restricting movement. The shape and the orientation of these small joints vary according to their location. The articulating surface of each superior and inferior articular process is known as the articular facet. The junction between the superior and inferior articular facets on one side of two adjacent vertebrae is known as a zygapophyseal joint (Z joint). Thus, there is a left and right Z joint between each pair of vertebrae.14 The planar joint surfaces are covered with hyaline cartilage approximately 1 mm to 3 mm in thickness that thins with age.14 Depending on their orientation, which varies along the length of the spine, the Z joints contribute to weight bearing. Axial load-sharing between anterior and posterior elements appears controversial with weight bearing by the Z joints dependent on spinal location and posture.
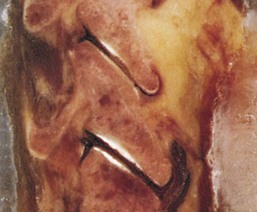
Uncovertebral Joints (Joints of Luschka)
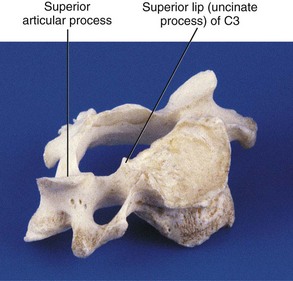
When viewed from the lateral or anterior aspect, raised lips at the superior aspect of the typical cervical vertebral bodies become apparent. These structures arise as elevations of the lateral and posterior rims on the top surface of the vertebral bodies (uncinate processes) (Figure 2-9). Normally the uncinate processes allow for flexion and extension of the cervical spine and help to limit lateral flexion. In addition, the uncinate processes serve as barriers to posterior and lateral intervertebral disc protrusion. When the uncinate processes of one vertebra articulate with the small indentation found on the inferior surface of the vertebra above, the articulations are referred to as uncovertebral joints or joints of Luschka. These small synovial joints are limited medially by the intervertebral discs and laterally by the capsular ligaments. The uncovertebral joints frequently undergo degeneration with resulting bony outgrowth. The degree of uncovertebral joint arthrosis parallels the loss of disc height. Osteophytes from the uncovertebral joint can potentially impinge upon the adjacent exiting cervical spinal nerve root and can deflect the course of the vertebral artery.7 Hall, in 1965, described three stages of degeneration of the joints of Luschka.15
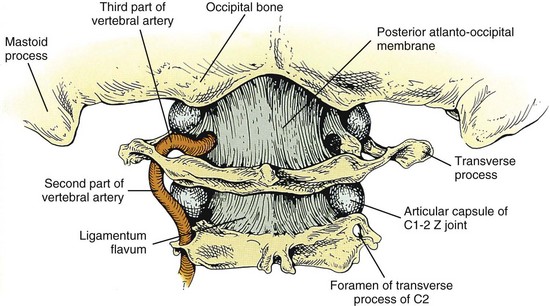
Spinal Ligaments
The Articular Capsules
The articular capsules of the Z joints are thin and loose and attached peripherally to the articular facets of adjacent zygapophyses7 (Figure 2-10). The capsules are longer and looser in the cervical region than in the lumbar and thoracic regions to accommodate for the greater amount of movement in the cervical region. The capsule consists of an outer layer of dense fibroelastic connective tissue, a vascular central layer made up of areolar tissue and loose connective tissue, and an inner layer made up of a synovial membrane.7 The anterior and medial aspect of the Z joint capsule is made up of the ligamentum flavum. The synovial membrane lines the articular capsule including the portion made up by the ligamentum flavum.7
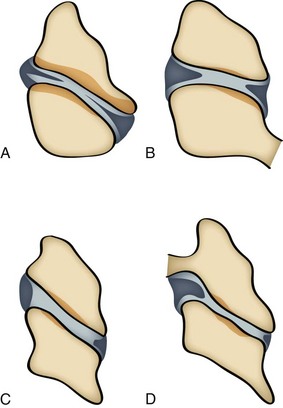
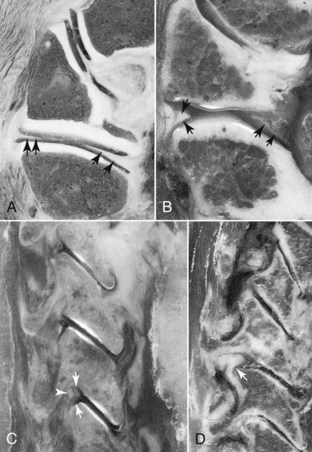
Synovial folds are synovial lined extensions of the capsule that protrude into the joint space covering part of the articular cartilage.16,17 These intracapsular structures have been identified as synovial fold projections into the Z joint at all levels of the cervical spine.18 Yu et al18 identified four distinct types of cervical Z joint menisci on magnetic resonance imaging. These menisci ranged from thin rims to thick protruding folds (Figure 2-11).
The ligaments of the spine are arranged so as to provide postural support between vertebrae, with a minimum expenditure of energy, while at the same time allowing for adequate spinal motion.2 They must also restrict motion within physiological limits so as to protect the neural elements of the spine. The spinal ligaments are designed to resist tensile forces acting parallel to the direction in which the fibers run. They also contribute to the maintenance of natural spinal curvature. Vertebral ligaments may be classified as long spinal ligaments and short intersegmental ligaments. The long spinal ligaments are continuous supporting bands running the entire length of the spine, while the intersegmental bands connect adjacent vertebrae.
Long Spinal Ligaments
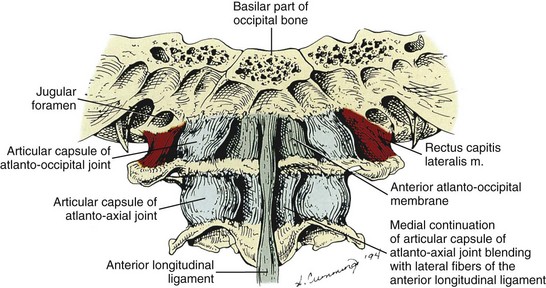
The long spinal ligaments include the anterior and posterior longitudinal ligaments and the supraspinous ligament. The anterior longitudinal ligament is a broad fibrous network attached to the anterior surfaces of the vertebral bodies. It forms a strong bond that runs from the anterior upper sacrum to the cervical spine where it attaches to the front of the body C2, the anterior tubercle of C1, and to the basilar occipital bone. The width of the anterior longitudinal ligament is diminished at the level of each intervertebral disc where it attaches to the annular fibers. It has several layers, the most superficial fibers being longest and extending over three to four vertebrae. The intermediate fibers extend between two and three vertebrae, while the deepest fibers run from one body to the next. Laterally short fibers connect adjacent vertebrae.6 The anterior longitudinal ligament narrows considerably between the atlas and occipital bone and blends with the atlanto-occipital membrane (Figure 2-12). Functionally, the anterior longitudinal ligament limits extension and excessive lordosis in the cervical region. It is frequently damaged in extension injuries to the cervical19 and lumbar areas.7
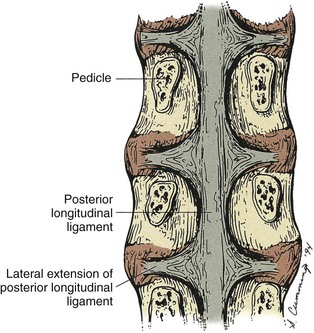
The posterior longitudinal ligament, unlike the anterior longitudinal ligament, is wider at the level of the intervertebral discs where it is interwoven into the annular fibers (Figure 2-13). It runs inside the spinal canal from the sacrum to the body of the axis. It attaches to the posterior superior and posterior inferior margins of the vertebral bodies. The upward extension of the posterior longitudinal ligament is the tectoral membrane, which continues cephalad as a strong broad band also attached to the basilar part of the occiput6 (Figure 2-14). The ligament is wide band in the cervical spine attaching to the intervertebral discs and adjacent vertebral bodies, narrowing and becoming more denticulate as it descends toward the lumbar region where it limits the ability to restrain posterior bulging of the intervertebral discs.20 Its superficial fibers bridge three or four vertebrae, while deeper fibers extend between adjacent vertebrae as perivertebral ligaments close to and fused with the intervertebral disc.6 A perivertebral membrane anterior to the deepest layer and attaching to the pedicles has also been found.20 The posterior longitudinal ligament adds stability to the vertebral column.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


