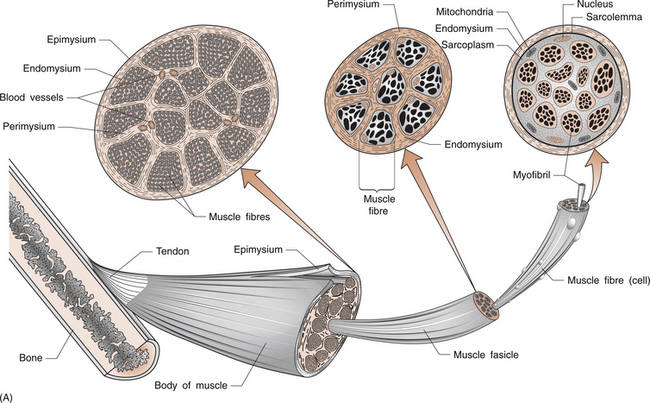
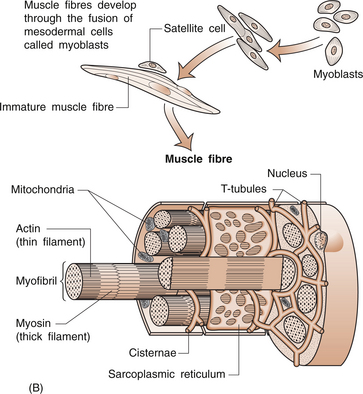
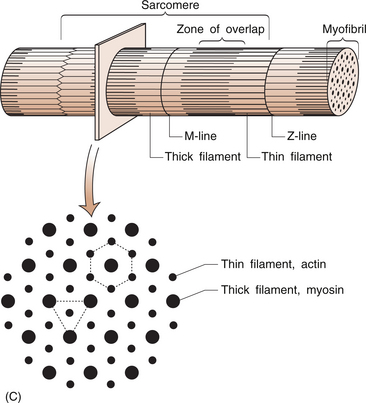
8 Rodger Laurent Muscles are made up of a collection of individual muscle fibres (Figs 8.1, 8.2). Each fibre is a multinucleated cell, which can be up to 10 cm in length with a diameter ranging from 10–100 μm. Normal muscles have the nuclei arranged around the periphery of the cells. The muscle fibre contains numerous myofibrils, which are 1–2 μm in diameter and the length of the cell (Fig. 8.1A). Myofibrils shorten and are the structures responsible for muscle contraction. They shorten the fibre because they are attached to the sarcolemma at each of its ends. Mitochondria and glycogen granules are situated between the myofibrils. The myofibrils consist of bundles of filaments, which are made up of the proteins actin and myosin, and are organized in repeating functional units called sarco-meres. Sarcomeres are the smallest functional units of the muscle fibre (Fig. 8.1C). The actin filaments are thin and the myosin filaments are thick. The thick filaments lie at the centre of the sarcomere with the thin filaments at either end. On either side of the centre, there is an area of overlap between the thin and thick filaments in which each myosin filament is surrounded by a hexagonal array of actin filaments. The arrangement of the myosin and actin filaments gives a banded appearance to the muscle and is the reason it is called striated muscle. The sarco-meres are separated by a dense area called the Z line. The M line is at the middle of the sarcomere and consists of proteins that bind the thick myosin filaments. The actin and myosin filaments are joined by molecular cross-bridges and, during contraction, these cross-bridges repeatedly disengage and engage at successive sites, with the result that the actin and myosin filaments slide upon one another and the myofibrils shorten. Each fibre is surrounded by a thin layer of collagen, called the endomysium. The fibres are then joined together in bundles to form fascicles, which are surrounded by a further layer of connective tissue called the perimysium (Fig. 8.1A). Groups of fascicles form the whole muscle, which is surrounded by a strong layer of collagen, called the epimysium. The epimysium merges with the peritenon of the tendon and the periosteum. The arrangement of the fascicles is variable and depends on the task of that specific muscle. Factors such as the amount and direction of the force required or the amount of muscle shortening determine the muscle architecture. Two examples are an arrangement of the fascicles parallel to the long axis of the muscle, as in the gastrocnemius (calf) muscle, or a convergent arrangement where the origin covers a wide area and the fascicles converge to a common attachment site, as in the pectoralis major muscle. The pectoralis major muscle has its origin covering a wide number of ribs, converging to a tendon that attaches to the upper humerus. Muscle fibres (Table 8.1) are divided on the basis of their morphology and physicochemical characteristics into two major groups: type 1 and type 2 fibres. Each type has different functions. The type 1 muscle fibres, slow oxidative, have a slow speed of contraction and a high resistance to fatigue. Their metabolism is oxidative and they have an increased concentration of myoglobin, which has an increased capacity to transport oxygen. They also have numerous mitochondria. Type 1 fibres generally have a greater capillary blood supply than type 2 fibres. A muscle spindle is a spindle-shaped stretch receptor found in most muscles but especially concentrated in muscles that exert fine motor control, such as the small muscles of the hand. The muscle spindle is about 100 μm in diameter and up to 10 mm in length. Muscle spindles receive a sensory innervation from groups Ia and II afferent nerve fibres and a motor supply from dynamic γ and static δ motor axons (see Ch. 3 for revision of nerve fibre types). All of the muscle fibres controlled by a single motor neuron form a motor unit. Small motor units where a motor neuron may control two or three muscle fibres are found in muscles where fine control is required. The converse is found in muscles that do not need fine control, for example the gastrocnemius or gluteus maximus. The amount of tension produced in a contracting muscle depends on the frequency of stimulation and the number of muscle units involved.
DISORDERS OF SKELETAL MUSCLE
Anatomy of skeletal muscle
The muscle fibre
Skeletal muscle structure
Types of muscle fibres
Muscle and tendon receptors
Control of muscle function
< div class='tao-gold-member'>
![]()
Stay updated, free articles. Join our Telegram channel








Full access? Get Clinical Tree


