Fig. 17.1
Chest sweat rate (chest SR) plotted against esophageal temperature (T es) for one subject exercising at an ambient temperature (T a) of 25 °C. The responses to increase chest SR to an increased Tes during exercise increased after a 10-day training in a temperate condition (T a, 20 °C) and further after a subsequent 10-day training in a hot condition (T a, 35 °C) [26]
17.4.1 Effects of PV Expansion on Thermoregulation
The exercise-induced PV expansion is suggested to be primarily associated with an increase in total volume of extracellular fluid [32] and secondary with an increase in plasma protein, mainly albumin content, resulting in drawing fluids into the intravascular space from the interstitium [32–34]. As for the mechanisms about the former, a facilitated Na+ and water reabsorption [32, 35] and also an increased voluntary fluid intake with an enhanced thirst sensation [36] due to an activated renin-angiotensin-aldosterone system and vasopressin release during and after exercise or dehydration are suggested [37]. As for the mechanisms about the latter, an enhanced plasma protein synthesis in the liver [38, 39] and an increased protein translocation from the interstitium to the intravascular space [40] with a decreased transcapillary protein escape ratio [41] are suggested.
Expanded PV after exercise training associates with an attenuated increase in Posm due to reduced lactic acid concentration in blood (enhanced lactate threshold) during exercise at the same absolute intensity and therefore shifts the core temperature threshold for cutaneous vasodilation and sweating downward. Moreover, expanded PV increases venous return to the heart and cardiac filling pressure and therefore enhances cardiac stroke volume and the response to increase SkBF and sweat rate to an increased core temperature during exercise [11, 16, 17]. Indeed, Ichinose et al. [27] suggested that an increase in cardiac stroke volume and also the sensitivity to increase SkBF in response to an increased Tes during exercise in a warm environment (Ta, 30 °C) were closely associated with an increased PV after 10-day endurance training (60 % VO2max for 1 h/day) in the same condition. More directly, Goto et al. [42] examined the effects of post-exercise protein and carbohydrate (CHO) mixture (Pro-CHO; 3.6 kcal and 0.36 g protein/kg) intake during 5-day exercise training (70 % VO2max for 30 min/day) in a warm environment (Ta, 30 °C) on PV and thermoregulatory responses. They suggested that plasma albumin content (Albcont) and therefore PV in the Pro-CHO group increased by ~10 % and ~8 %, respectively, which were significantly higher than ~4 % in the placebo intake group (CNT; 0.9 kcal and 0 g protein/kg body weight). They attributed that the increase in Albcont was caused by an enhanced hepatic albumin synthesis after exercise because of an enhanced bioavailability of the substance [38, 39] and anabolic effects of insulin on hepatocytes [43]. Most importantly, they found that the sensitivity to increase SkBF and sweat rate in response to an increased Tes in the Pro-CHO group increased by 44 % and 56 %, respectively, after training, which were significantly higher than those in the CNT group increased by 10 % and 19 %, respectively (Fig. 17.2). Moreover, the increases in heart rate and Tes during exercise were attenuated after training in both groups but more prominent in the Pro-CHO group than in the CNT group, indicating decreased cardiovascular and thermal strains after training with PV expansion (Fig. 17.3).
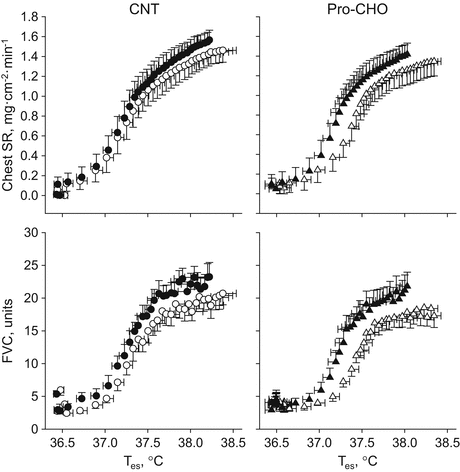
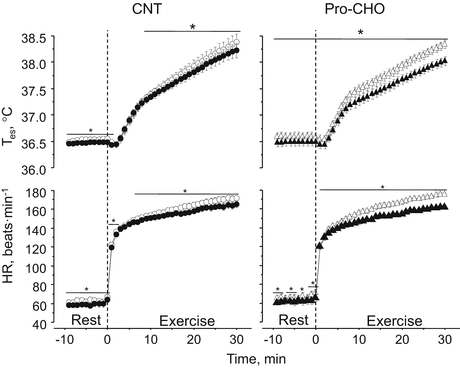
Fig. 17.2
Chest sweat rate (chest SR) (a, b) and forearm vascular conductance (FVC) (c, d) responses to increased esophageal temperature (T es) during exercise in a warm environment (T a, 30 °C) (open symbols) and after (closed symbols) 5-day training in the same condition. CNT, placebo intake group; Pro-CHO, protein and carbohydrate intake group. Means and SE bars are presented for nine subjects [42]
Fig. 17.3
Esophageal temperature (T es) and heart rate (HR) responses during exercise in a warm environment (T a, 30 °C) before (open symbols) and after (closed symbols) 5-day training. CNT, placebo intake group; Pro-CHO, protein and carbohydrate intake group. Means and SE bars are presented for nine subjects. *significant differences vs. before training, P < 0.05 [42]
In addition, Ikegawa et al. [30] confirmed the observations by Goto et al. [42] by showing an increased PV with a downward-shifted Tes threshold for cutaneous vasodilation and sweating after the same training protocol and further the downward-shifted Tes threshold were totally or at least in part abolished after the increased PV was normalized by administration of diuretics after training. Furthermore, Ichinose et al. [27] suggested that the sensitivity of the upward shift of Tes threshold for cutaneous vasodilation at a given increase in Posm by hypertonic saline infusion was reduced after 10-day exercise training (60 % VO2max for 60 min/day) in a warm environment (Ta, 30 °C) although that for sweating remained unchanged. They further suggested that the reduction in each subject was significantly correlated with the increase in PV after training, suggesting that the stretch of cardiopulmonary baroreceptors due to PV expansion was associated with the reduction. These results clearly demonstrate that the enhanced thermoregulatory responses and cardiovascular capacities after exercise training are closely associated with PV expansion in addition to the neural adaptation of the thermoregulatory center and thermoregulatory effectors [1, 12].
17.4.2 Effects of Other Adaptations with Training on Thermoregulation
In addition, Na+ concentration of sweat decreased after exercise training or heat acclimation due to an enhanced Na+ reabsorption at the sweat gland duct with an enhanced sensitivity to aldosterone [44]. Hypotonic sweat is advantageous for maintaining PV during exercise because hypotonic sweat loss causes a greater increase in osmolality of extracellular fluid that facilitates water movement from intracellular fluid to extracellular fluid space according to osmotic gradient between the spaces [44] and therefore attenuates the reduction in venous return to the heart. As a result, the decrease in thermoregulatory responses with a given loss of sweat is attenuated with low sweat Na+ concentration [45]. Moreover, the upward shift of Tes thresholds for cutaneous vasodilation and sweating at a given increase in Posm was reduced in heat-acclimated subjects in whom an increase in Posm at a given loss of sweat was enhanced due to low sweat Na+ concentration [45].
17.5 Effects of Biological Aging on Thermoregulation
The susceptibility to heat-related illnesses in the elderly [6] is caused by deteriorated thermoregulatory responses with aging [46]. Actually, previous studies have indicated that healthy elderly individuals have an impaired thermal perception [47] and impaired autonomic [48, 49] and behavioral [50, 51] thermoregulatory responses. Decreased VO2max and cardiovascular capacity are associated with the deteriorated thermoregulatory responses in the elderly [49]. However, it has been suggested that elderly subjects with matched VO2max to younger subjects show a lowered SkBF response to total body or local passive heating as well as during exercise in a hot environment than younger subjects [52, 53]. Kenney et al. [53] suggested that the limited SkBF response to hyperthermia during exercise in a hot condition was caused not by an enhanced vasoconstrictor system but mainly by a decreased sensitivity of active vasodilator system to increased Tes, by using bretylium tosylate to block local release of norepinephrine on the forearm skin. In addition, total body and local sweat rate in response to passive heating or exercise are lower in the elderly than in young adults [54].
Moreover, decreased PV and increased Posm at baseline condition [55], diminished thirst sensation [55] and responses in antidiuretic hormone and aldosterone after thermal dehydration [37], decreased renal concentrating ability [56], and lower reabsorptive ability of sweat gland ducts [56] with advancing age are suggested to be associated with the decreased thermoregulatory responses in the elderly. Splanchnic and renal vasoconstriction during exercise, which enhance the redistribution of cardiac output to the skin vasculatures and associate with an increased cutaneous vasodilation in the youth, were also decreased in the elderly [57]. Furthermore, it is not uncommon for the elderly to be taking a variety of prescription drugs that may affect thermoregulatory responses and body fluid regulation [1]. These diminished body fluid regulations and thermoregulatory functions with aging can be improved with exercise training and heat acclimation, although generally the gain is lower or limited relative to their younger counterparts.
17.5.1 Effects of Exercise Training on Thermoregulation in the Elderly
Several longitudinal studies have suggested that thermoregulatory responses during exercise in a hot environment are enhanced with aerobic training performed under cool, thermoneutral, or hot conditions in initially sedentary elderly subjects [29, 57, 58] as well as in young subjects [26–28]. Thomas et al. [29] reported that a 16-week aerobic training lowered the mean body temperature threshold for cutaneous vasodilation during exercise in a hot environment (Ta, 36 °C) both in young and elderly subjects, whose VO2max increased by ≥5 %. They also suggested that the enhanced cutaneous vasodilation is caused by an enhanced sensitivity of active vasodilator system [29]. Additionally, Okazaki et al. [58] confirmed and extended these results by showing that Tes thresholds for cutaneous vasodilation and sweating during exercise in a warm environment (T a, 30 °C) were lowered after an 18-week aerobic and resistance training under cool and thermoneutral conditions, with improved VO2max, by 20 % and 10 %, respectively, in initially sedentary elderly subjects. In both studies, on the other hand, the sensitivities to increase SkBF and sweat rate in response to an increased mean body temperature or Tes remained unchanged after training in elderly subjects. Okazaki et al. [58] attributed the results to an attenuated PV expansion after training in elderly subjects, by showing a linear correlation between changes in the sensitivities and those in PV after training. As above, in young subjects, PV expansion is considered to be a major mechanism involved in the training-induced enhancement of thermoregulatory responses by increasing cardiac stroke volume and/or by suppressing baroreflex-induced attenuation of skin vasodilation by increasing venous return to the heart [11]. Additionally, Ho et al. [57] showed that splanchnic and renal vasoconstriction during exercise were enhanced in young subjects but not in elderly subjects after aerobic training. Thus, the beneficial effects of training on thermoregulation in elderly people involve an increased VO2max but are generally attenuated compared with younger counterparts [57, 58].
The blunted increase in PV with aerobic training in elderly people is likely to be associated with a reduced fluid intake after thermal dehydration [37, 55] or water deprivation [59] with aging; however, an attenuated increase in Albcont with exercise [34, 58, 60] appears to be a primary limiting factor. As above, one mechanism of the increased Albcont after aerobic training is likely to be an enhanced hepatic albumin synthesis response to exercise in the younger group [38, 39]; the smaller increase in Albcont after training in the elderly group could be caused, in part, by the blunted response of the albumin synthesis to exercise [61, 62] due to reduced gene expression with aging. However, it is also plausible that this is caused by protein intake insufficient for albumin synthesis in elderly people since they are likely habituated to low energy and protein diets due to minimal daily physical activity, and therefore it would be improved if substrates for plasma albumin synthesis are given immediately after exercise where albumin synthesis is reportedly enhanced [39, 62]. In this regard, one study showed that recoveries in Albcont and PV after acute high-intensity interval exercise were generally attenuated in elderly versus young subjects when they consumed a non-energy placebo just after exercise, whereas Albcont and PV recovered more when they consumed a protein and CHO mixture than when they consumed the placebo in elderly and young subjects [34].
< div class='tao-gold-member'>
Only gold members can continue reading. Log In or Register to continue
Related posts:
 Bone Disease Associated with Diabetes Mellitus: Particularly Focusing on Its Contribution to the Development of Atherosclerosis
Mechanism of Skeletal Muscle Contraction: Intracellular Signaling in Skeletal Muscle Contraction
Bone Disease Associated with Diabetes Mellitus: Particularly Focusing on Its Contribution to the Development of Atherosclerosis
Mechanism of Skeletal Muscle Contraction: Intracellular Signaling in Skeletal Muscle Contraction
 Mechanism of Skeletal Muscle Contraction: Role of Mechanical Muscle Contraction in Glucose Homeostasis
Mechanism of Skeletal Muscle Contraction: Role of Mechanical Muscle Contraction in Glucose Homeostasis
 Ectopic Fat Accumulation and Glucose Homeostasis: Role of Leptin in Glucose and Lipid Metabolism and Mass Maintenance in Skeletal Muscle
Ectopic Fat Accumulation and Glucose Homeostasis: Role of Leptin in Glucose and Lipid Metabolism and Mass Maintenance in Skeletal Muscle
 Ectopic Fat Accumulation and Glucose Homeostasis: Ectopic Fat Accumulation in Muscle
Ectopic Fat Accumulation and Glucose Homeostasis: Ectopic Fat Accumulation in Muscle
 Fracture Risk in Diabetes
Fracture Risk in Diabetes
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


